顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
獲得免疫の2つの道
2015年1月15日
前回紹介した有顎類進化に関するおとぎ話が決定的にまちがっている点が一つある。それは免疫系の進化についてのくだりだ。この話では、美食家になった私たち先祖が、食物を通した感染から身を守るため免疫系を発達させたと紹介したが、実際には現存の八つ目ウナギやヌタウナギにも立派な免疫システムが存在していることがわかっている。八つ目ウナギの免疫システムを担う分子を仔細に眺めると、分子としては有顎類の抗原結合分子と全く違っているのに、免疫系全体でみると私たち顎のある脊椎動物と多くの共通性を持っている。両者の比較は、進化過程で新しい機能が生まれる過程での共通性と多様性について様々な示唆を与えてくれる。今回はゲノム研究から明らかになった無額類の免疫システムと私たちの免疫システムを比べてみよう。
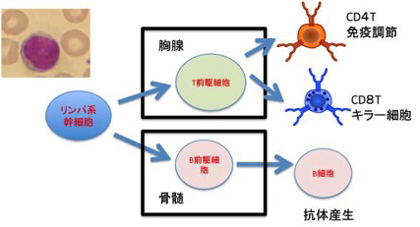
図1 左上が末梢血中のリンパ球。リンパ球は骨髄で作られ抗体を作る細胞に分化するB細胞と、胸腺で作られ免疫調節に関わるCD4T細胞と、細胞などを殺す役割を持つCD8T細胞に分かれる。これ以外に、胸腺を経ずに分化するγδT細胞が存在する。
まず私たち人間も属している有顎類の免疫系について簡単にまとめておこう。免疫反応はリンパ球と呼ばれる細胞質の少ない細胞によって担われている(図1)。さらにリンパ球はB細胞とT細胞に別れる。B細胞は抗原と特異的に結合する抗体(Ab)産生を造る細胞で、発生過程で遺伝子の再構成を行うことで、特定の抗原だけに反応する抗体を造る細胞になる(図1及び、第10話参照)。一方胸腺に移動したリンパ球前駆細胞は、抗体遺伝子とは別の抗原反応分子T細胞受容体 (TcR)遺伝子再構成を行い、抗原特異的T細胞になる。膨大なレパートリーを持つ抗原特異的T、B細胞が、遺伝子再構成や突然変異を通して形成される過程についてはすでに第10話で述べたので参考にしてほしい。重要なのは、1)多様な抗原に対応するために数多くの抗体V遺伝子が存在すること、2)遺伝子の発現には離れている遺伝子(VとD、DとJ)を選択する遺伝子再構成過程が必要で、3)これにより単一細胞が1抗体遺伝子(or TcR)だけを発現し、一種類の抗原特異性だけを持つという条件が達成されるとともに、4)遺伝子結合部位にランダムな変異が入ることで、抗体のレパートリーが増大する、の4点だ。この過程はAbもTcRも同じで、遺伝子再構成に関わる分子も同じだ(Rag1,Rag2)。さらに、分子構造的にはAbもTcRも免疫グロブリンスーパーファミリーに属しており、進化的にも共通の祖先に由来する。
一方、八つ目ウナギやヌタウナギの仲間の血中にリンパ球に似た細胞は確かに存在する。しかし、T細胞を作る胸腺は存在せず、またAbやTcRと相同な免疫グロブリンスーパーファミリーに属する抗原結合分子は見つからなかった。このため22話で紹介したおとぎ話のように、Ab, TcR, 胸腺は下顎の進化と並行して新たに発生したと考えられてきた。ところが2004年に、ロイシンを多く含む繰り返し構造を持った、しかも多様性を持つcDNAが八つ目ウナギから分離され、LRR(leucin rich repeat 分子)と名付けられた。Leucin rich repeat構造は無脊椎動物も含む多くの種に分布する自然免疫に関わるTLR受容体に存在する。しかし、TLR遺伝子の数は10種類を超えているが、一つのTLR分子の中での多様性は全くない。一方、LRR遺伝子構造の研究かから、LRRがAbやTcR分子と同じで、多様性のない定常部分と、配列に大きな多様性が存在する可変部分からできており、成熟個体から分離されるLRRの遺伝子配列はクローンごとに違うことがわかり、可変性リンパ球受容体(variable lymphocyte receptor:VLR)と名付けられた(図2)。その後、この抗原結合部分子にはVLRA,VLRB,VLRCの3種類が存在し、この内のVLRBは抗体と同じで血中に分泌される構造を持っていることが明らかになった。この2つの種は、多様な抗原に反応するという同じ機能を、分子構造が全く異なる(即ち起源の全く異なる)祖先分子から進化させてきたことがわかる。
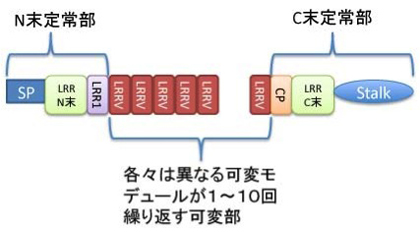
図2 無学類抗原結合分子可変性リンパ球受容体(VLR)の分子構造。ほとんどのドメインがLeucin rich repeat構造を持っており、中央にそれぞれのモデュールの遺伝子配列が異なる、LRRVが繰り返すことで可変部を形成する。LRRVがいくつ存在しているかによって分子の大きさは異なる。
実は私たち有顎類の抗原反応分子も3種類、即ち抗体分子と、αβTcR、γδTcRの3種類が存在する。偶然とはいえ、3種類の抗原結合分子がセットになって存在しているのを見ると、免疫系を構築するための必要条件がなにかありそうだ。
無顎類に独立の抗原反応分子があることは明らかになったが、ではどの細胞がこれらの抗原反応分子を発現しているだろうか?リンパ球はあるのだろうか?有額類のT、B細胞のように区別があって、異なる組織で発生するのだろうか?無顎類にも胸腺に似た組織はあるのだろうか?LRR分子の発見とともに多くの疑問が残った。幸いLRR発見後の10年間で様々なことがわかり、詳細は別にして無顎類免疫系の大きな枠組みは理解できるようになっている。
まず、無顎類にもリンパ球が存在する。またそれぞれのリンパ球は3種類のVLR分子のうちどれか一つだけ発現しており、しかも有顎類のリンパ球と同じで、2つある対立遺伝子の一方だけが発現していることが明らかになった。このことは、発現している抗原結合分子の種類に応じて3種類のリンパ球が存在することを意味している。即ち、VLRBを分泌するリンパ球が有顎類のB細胞に対応し、VLRAとVLRCを発現するリンパ球がT細胞に対応することになる。ではこれらの細胞はどこで作られるのか?特に、胸腺がない無顎類でT細胞に対応するVLRA発現細胞はどこで作られるのかが調べられた。八つ目ウナギの幼生時期の様々な組織でVLRB分子の発現を調べると、VLRB陽性細胞は腎臓や、typhlosoleと呼ばれる腸管内の造血組織に存在していた。これは有顎類のB細胞が様々な造血組織で作られるのと同じだ。一方驚くべきことに、T細胞に当たるVLRA陽性細胞はエラの先端部に限局して存在していた。「驚くべきことに」と書いたのは、有顎類の胸腺は鰓弓と呼ばれるエラに分化することが運命づけられた胎児組織から発生するからだ。胸腺が臓器として進化する前から鰓弓はT細胞を発生させる能力を持っていたことになる。脊椎動物になって発生したリンパ球をT,B細胞に2分して異なる機能を担わせるとともに、それぞれを別の組織で分化させる細胞システムは、脊椎動物の進化後比較的早期に完成していたようだ。一方、抗原に対応するためそれぞれの種が発展させた分子は異なる遺伝子を起源にしていることは明らかなので、無顎類と有顎類は2種類のリンパ球を使う体制が完成したあと、独自に抗原反応分子を分化させたと考えられる。
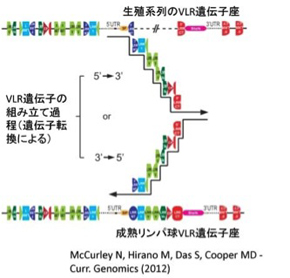
図3 リンパ球の分化に伴い、VLR遺伝子の定常部の間に、可変部が集められていく。この過程は遺伝子転換によると考えられており、3'側からも、5'側からも起こる
最後にVLRの多様性がどのように発生するのかについて現在明らかになっていることを紹介しよう。VLRの多様性がどのように生まれるのかを調べるため、ゲノムの構造が調べられ、予想通りというべきか、驚くべき一致というべきか、無顎類のリンパ球への分化の過程でもVLR分子の定常部遺伝子の間に可変部LRRVモデュールが集められることがわかった。この過程について、この分野をリードするマックス・クーパー(Max Cooper)らがオープンアクセス(誰もが無料で閲覧可能な状態)で提供してくれている図3を使って紹介しよう。まずリンパ球分化が始まる前のゲノム構造は無顎類の方がずっと複雑だ。VLR分子5'及び3'端分子をコードする定常ドメイン遺伝子の外に、可変部を構成するLRRV遺伝子が散らばって存在している(VLRB遺伝子では約700Kbのなかに820の異なるLRRVが存在している)。リンパ球分化前のゲノムでは5'及び3'両端の定常ドメインの間にはイントロンが存在しており、この遺伝子構造は発現することができない。リンパ球分化が始まると、このイントロンがゲノム上に散らばっているLRRVモデュールと置き換わっていく(図3)。この過程を通して、定常ドメインの遺伝子は同じ場所で変化することなく存在したまま、イントロンだけがLRRVに置き換わっていく。したがって、遺伝子の一部が欠損する遺伝子再構成が起こっていることは考えられない。完全に証明されたわけではないが、おそらく段階的な遺伝子転換(gene conversion)で可変ドメインが定常ドメインの間に集められるのではと考えられている。この過程の途中段階のリンパ球のゲノムから、遺伝子再構成がないことが確認され、また遺伝子転換によるイントロンと可変モデュールの置き換えが5'側からも3'側からも起こることがわかってきた(図3)。実際、遺伝子再構成に必要なRag1,Rag2遺伝子は無顎類には存在しない。代わりに、有顎類の免疫グロルリンクラススウィッチを調節するAID分子と同じファミリーに属するCAD1,CAD2という分子が関わることもわかってきた。詳細は省くが、遺伝子転換により可変モデュールを集めることで、例えば無顎類のVLRBは1014〜1017という膨大な数のレパートリーを形成することができる。また、血中に存在する抗原特異性がわかっているVLRBの抗原との結合の強さは、私たちの抗体に十分匹敵するようだ。このように無顎類は美味しいものを食べることはできなかったが、外界からの侵入に対応するため、有顎類とは全く別の多様性のある抗原結合分子を発達させ、免疫システム進化のもう一つの可能性を示したと言える。
最後に以上をまとめた無顎類と、有顎類の免疫系の進化の道筋を図4に示しておく。
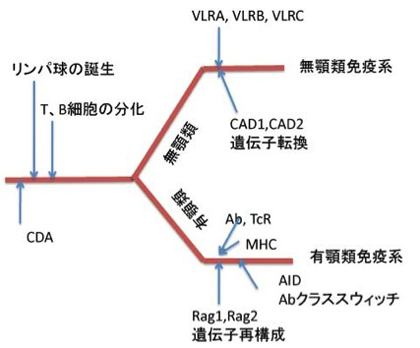
図4 脊椎動物が発生して、T、Bリンパ球体制が先に完成し、この後別々の抗原反応分子が開発されたことを示している。最初のT、B細胞がどの抗原反応分子を使っていたのかはわからない。ただ、遺伝子再構成は有顎類の進化とともに始まったと考えられる。
今回は少し難しかったかもしれない。ただ免疫システムの進化は、新しい機能が進化過程から生まれる時の多様性と、相同性の両方を知るための格好の例になっているのを楽しんでほしい。
この稿ではThomas Boehm等の総説論文、Boehme et al, VLR-based adaptive immuneity, Annu. Rev. Immunol. 30:203, 2012を参考にした。