顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
自然選択とクローン選択
2014年7月1日

これまで内部共生や水平遺伝子伝搬と言った大きなゲノムの変化について紹介して来た。実際、真核生物の進化と言った大きな変化を理解するためには、多くの遺伝子が一度に獲得される機構の解明は重要だ。ただゲノムの大きな変化は生物としての独自性を揺るがし安定性を損なうため、ゲノム多様性の獲得機構としていつも起こるわけではない。ではゲノム多様化はどのようなメカニズムで起こっているのだろうか?この問いはもちろん進化研究の課題だが、実は免疫学も同じ課題を共有している。免疫学の歴史はジェンナーの種痘から始まる。ただ、私たちが多様な外来の病原体に対して抵抗する免疫反応を持っていることを理解するようになったのは、1)ニワトリコレラ、炭疽病、そして有名な狂犬病ワクチン開発に成功したパスツール、2)ワクチンの効果が血清中の特異的反応分子(抗体)の作用であることを証明したベーリングと北里、そして3)血液型を発見するとともに、単純な化学物質に対して特異的な抗体が出来ることを示したランドシュタイナーらの研究に負う所が大きい(図1)。

図1 免疫学を始めた偉人 ジェンナーからランドシュタイナーまで。
特にランドシュタイナーにより、ハプテンと呼ばれる単純化学物質に特異的に反応する抗体が出来ることが示され、更にこの化学物質の側鎖の小さな化学的違いすら区別する抗体が出来ることが証明されると、ほぼ無限の外来物質に対して抗体を作ることができると考えられるようになった。しかし一体どうしたらほぼ無限とも言える外来物質を区別する抗体ができるのか?この問いにチャレンジしたのが免疫学だが、この歴史を振り返ると、19世紀進化論で行われた議論が繰りかえされているのを見ることが出来る。ダーウィンの進化論の必須条件、多様性の獲得(Generator of diversity:GODとは意味深だ)機構に関しては、免疫反応から学ぶ方が理解し易いと思う。今回は趣向を変えて免疫理論での「GOD」について見てみたい。しかしその前に、19世紀の進化理論をおさらいしてみよう(図2)。

図2 ラマルクとダーウィンの進化論
生物が天地創造時から変わらず現在まで存在して来たとするキリスト教の教義が後退し、単純な生物が徐々に多様化、複雑化すると考える進化思想が生まれたのは19世紀だ。進化思想をはっきりと表明したのはダーウィンの祖父エラスムス・ダーウィンで、この考え方からラマルクの進化論が発展して来たと言える。図2にラマルクと、ダーウィンの進化論の比較をわかりやすく図示した。ラマルクは、高い所にある葉を食べると言う目的に会わせて身体が変化し、獲得した性質が子孫に伝わると考えた。一方、ダーウィンは、目的に会わせて身体が変わるといった都合のいいことは起こらず、高い所の葉を食べることが出来る首の長いキリンが最初から集団の中に存在し、選ばれて子孫を多く残すことで首の長いキリンだけになると考えた。明らかにラマルクの考えには目的へと進む生命の力の様なものが想定されている。ビュフォンを始め18世紀の自然史は様々な形でこの様な生命の力を想定してきた。一方ダーウィンの考え方からはこの様な生命の力が排除されていることに注目して欲しい。
さて免疫学は第二次大戦後大きく進展する。まず「ほとんど無限とも言える抗原に対して特異的に抗体が作れるのか?」という問いに対して2つの仮説が出された。一つは鋳型説で、もう一つがクローン選択説だった。先ず鋳型説を見てみよう(図3)。

図3 ポーリングが考えた抗原特異的抗体の産生(指令説)
この図はノーベル化学賞に輝いたポーリングによって1940年描かれたもので、同じアミノ酸配列を持つ抗体ポリペプチドが、抗原に出会うことで抗原に合わせて折り畳まれ、ほぼ無限ともいえる異なる立体構造抗体を作ることが出来ることを表現している。はてどこかで聞いたことがある話だ。高い木の枝に茂っている葉っぱを食べるために首が伸びるとしたラマルクの進化論に似ていないだろうか?首が伸びるために必要な生命の力に代わって、抗体自体が持つ自由に姿を変える力が仮定されている。一見分子論的な合理性があるように見えるが一種の目的論で、実際にはそんな都合のいい分子はない。私が免疫学に初めて出会ったときはポーリングの指令説は既に否定され、それに代わってオーストラリアのノーベル生物学賞受賞に輝いたバーネット(Burnet)博士が提唱していたクローン選択説が主流になっていた(図4)。
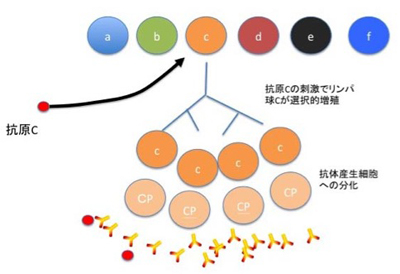
図4 バーネットが考えたクローン選択説
この図はバーネットが自分の説を説明するため書いた総説にある図を参考に作成し直した図だが、多様なリンパ球のうち抗原にヒットする細胞だけが増殖し、抗体を作る様が描かれている。まさに首の長いキリンが選択されるダーウィン進化論と同じ図だ。この図からわかるように、抗原に対する特異抗体は目的ではなく、最初から集団の多様性として存在しており、抗原はそれを選んで増幅するだけだ。20世紀後半に起こったこの論争を見ると、ダーウィンの進化論が受け入れられた後でも、私たちの頭から目的論的思考を追い出すことがいかに難しいかを思い知る。このように多様化と選択の枠組みで様々な歴史を説明する考えは、今や進化論や免疫学にとどまらず人文科学領域にも拡がっている。
クローン選択説が正しいとなると、私たちの身体の中には多様な抗原に匹敵する種類の抗体に対応する多様なB細胞が存在して、抗原が入ってくるとその中から抗原に適合したクローンが選ばれ、増殖し、抗体を作ることになる。ついに最初の問いに戻って来た!「ほぼ無限に近い抗原に匹敵する抗体の多様性が生み出すメカニズムは何か?」1970年代から多くの研究が行われ様々な説が提唱されたが、全ゲノム解析が多くの動物で終わった結果、様々な多様化の機構がフルに利用されていると言う結論が得られている。抗体の多様化の機構についてほ乳動物に限って見て行こう。本来なら抗体の分子構造から遺伝子に至るまで、詳しく述べる必要があるが、ここではGOD:多様性がどう生まれるかだけを問題にするので、抗体の中でもH鎖と呼ばれているペプチドをコードする遺伝子に話を限定する。
先ず私たち人間を見てみよう。子孫に伝えることの出来る生殖系列の遺伝子には80前後のVH遺伝子、23D遺伝子、16J遺伝子が離れて存在している。抗体を造るB細胞ができる時、これら遺伝子の中からVH,D,J各遺伝子が遺伝子再構成により一つに集められ、一つの機能的遺伝子だけが一個のB細胞で発現するようになる(図5)。
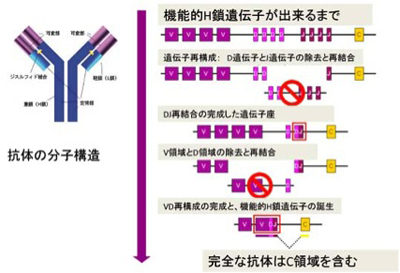
図5:抗体分子の構造と、遺伝子再構成による多様性
理論的には、この遺伝子再構成によって最低80x23x16(29440)、ほぼ3万種類の多様性を発生させられる。更に実際には、3つの遺伝子領域が再構成する際、それぞれの結合部位に核酸の欠損、挿入などが起こるため、多様性が少なく見積もっても100倍近く跳ね上がる。これだけでもう既に百万を超すが、更に同じ過程が抗体のもう一方のパートナー軽鎖遺伝子でも起こる。両者を掛け合わすとこれだけでほぼ無限のレパートリーが形成されると十分言えるだけの種類の抗体を作る事が出来る。比較的ヒトやサルに近いマウスでも全く同じ様に多様性が生まれるのが確認されている。即ち、先ず進化過程で遺伝子重複が起こり、遺伝可能な多様性が用意される。次にBリンパ球が分化する時に、遺伝子再構成、再構成時の核酸の欠損や添加、最後に通常の突然変異を積み重ねてほぼ無限の抗原に対応している。遺伝子の重複から、組み換え、突然変異まで全て揃っているのは進化的にヒトに近い種に限られるが、例えば牛のようにVH遺伝子を一個しか使わないほ乳動物もいる。この場合、再構成による変異と、点突然変異だけでほぼ無限の抗原に対応している。本当に大丈夫か?と問われそうだ。しかし今でも抗体を作るためによく使われるウサギも機能しうるV遺伝子を50近く持つにも関わらず、実際に再構成されているのは3−4種類だけで、それも90%以上はVh1と呼ばれる遺伝子だけだ。Dが11個、Jが6個あるので、100種類位の多様性は始めからあるが、マウスやヒトと比べるとずいぶんハンデがある。それでもほぼどんな抗原にも対応して抗体ができる。事実今でもウサギは抗体を造るために最も良く用いられる動物だ。実際には、V遺伝子間でおこる遺伝子変換と、点突然変異により多様性を発生させており、この様なメカニズムで十分ほぼ無限とも言える抗原に対応できるようだ。おそらく種の進化に必要な種内の多様性発生も、ほぼ同じメカニズムで起こるのだろう。まとめると、1)遺伝子重複、2)遺伝子組み換え時の遺伝子欠損や挿入、3)遺伝子変換などの組み換えメカニズム、4)点突然変異の組み合わせが、多様性の発生メカニズム、即ちGOD(generator of diversity)の正体だ。
※図1、図2、図5はWikimedia Commonsより。