LECTURE & TALK
私のきのこ学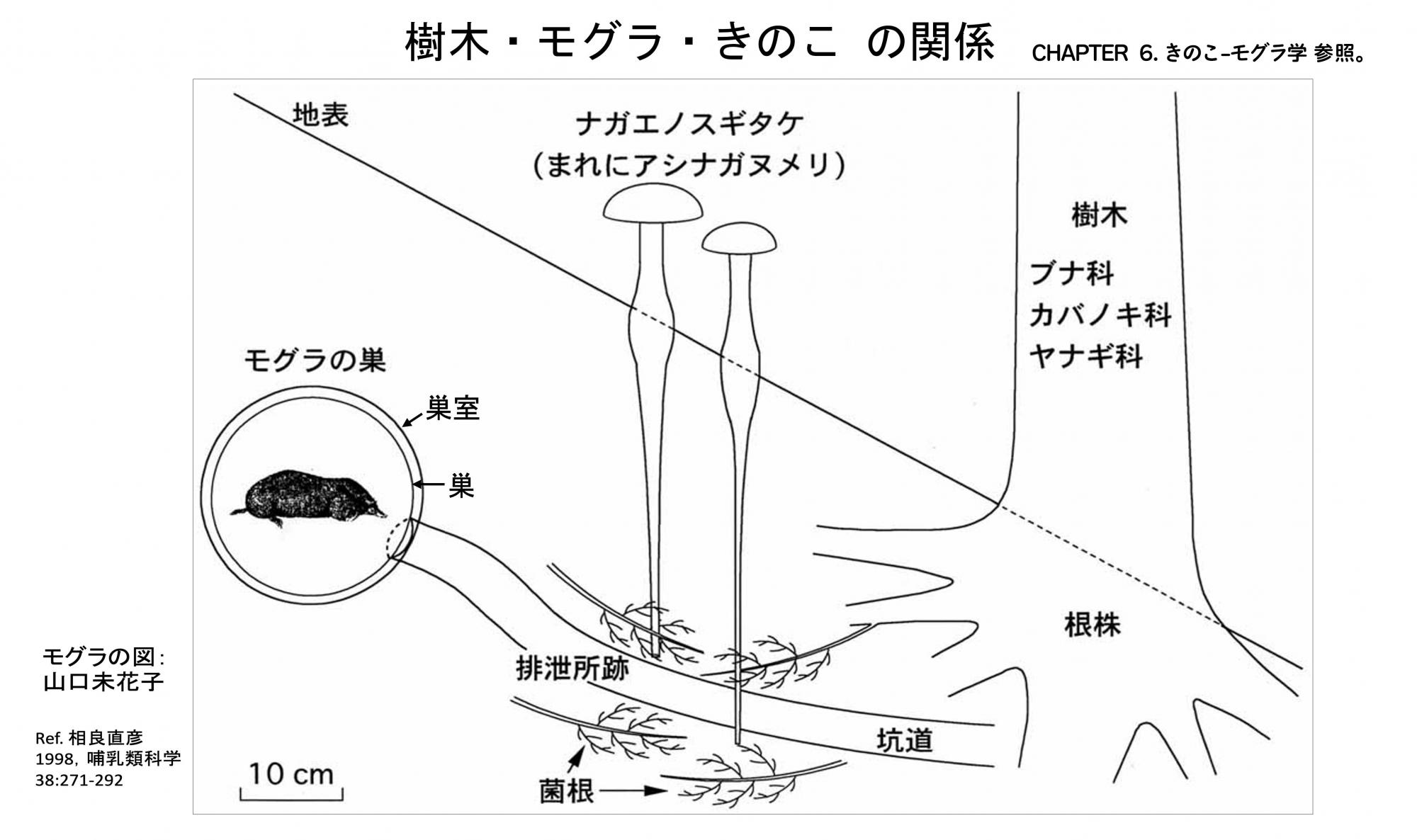
LECTURE & TALK
私のきのこ学講演

1. はじめに
九州の田舎から出て参りました。自己紹介として、2つお話しします。
京都大学農学部の私たちの学年は確か150名で入学して、3年生で各専門学科に分属する時、私の志望学科は定員28名に対して30人の志望者があり、試験で落とされた2人のうちの1人が私です。挫折感はほとんどありませんでした。そこでもし第1志望に行っていたら、人生は違ったものになって、きのこの研究はしなかったと思います。
現在は、まず、半農半学。この「農」は現代的な意味の農「業」でなく、昔風の、何でもやる百姓です。「学」の方は、もう現場の仕事はしておりません。そして昨年、2012年以来3度目の大洪水があり、甚大な土地被害を受けました。「二兎を追うものは一兎をも得ず」と申しますが、半農半学で二兎を追い、洪水対応ほか、三兎、四兎を追って死にそうです。


2. きのこについて-私の理解
菌類学をmycologyマイコロジーと言いますが、今日の演題の「私のきのこ学」をちょっと気取って言うと“ My mycology ” です。副題の、森における「後始末」と言えば、落葉落枝や枯木の後始末が大きな軸になりますが、私は、動物の排泄物や死体の行く末を取り上げます。ただし、初めから排泄物や死体を問題視していたわけではありません。
まず、きのこについて私が理解しているところを、ナガエノスギタケを例にお話しします。スライドをご覧ください(スライド1)。いわゆる「きのこ」です。これは「子実体」と言い、傘の裏側のひだで胞子をつくって飛散させるための構造物です。きのこの生活環で一時的に観察されますが、この姿できのこの生活が完結しているわけではありません。

(スライド1)
次に生活者としてのきのこを見ていきます(スライド2)。左の写真の地下部は「根」ではありません。「地中柄」と言います。地上部分と地中柄と全部あわせて一つのきのこ(子実体)です。子実体の一部分を切り取って試験管内の寒天培地で培養すると、まず生活の本体としての栄養菌糸が増殖します(写真中央)。菌類は我々と同じく光合成をしません。何かを食べなくてはなりません。我々は口から食べますが、菌類は細い菌糸の表面から養分を吸収して増殖します。ここでは、寒天に溶かしてある養分を吸収して増殖しています。栄養菌糸にエネルギーを蓄えて、温度などの条件が整うと子実体を形成します。

(スライド2)
昆虫は成虫になる時、幼虫の姿からは想像できないような姿に変態します。きのこもそのように見たらいい。私は、栄養菌糸から子実体への変化を、「変態」と理解しています。栄養菌糸の外見はどれも似たり寄ったりで種類は判別できませんが、子実体は千差万別です。ついでに見ていただきたいのは、試験管内にできた子実体には長い柄がありません。ナガエノスギタケの長い柄は、子実体の形態として必須ではないことがわかります。
栄養菌糸から子実体に「変態」する時、大きな子実体ができるには元が大きくなければならないし、きのこ(子実体)が沢山生えてきたら地中にそれだけの元があったからです。この原理を、私は我流に「質量保存の法則」と呼んでいます。きのこは霞を食って生えることはできません。
試験管内では、菌糸は寒天に溶けている養分を吸収して生活していますが、森の中ではモグラの排泄物由来の物質と植物の根から供給される養分で暮らしています。その時、菌糸は木の根と共生します。右上の写真で、白く見えるのは菌糸、黄色味を帯びて網目状に見えるものが根です。その根を薄く輪切りにしてみます(写真右下)。根の外側で緑色に染まって見えるのが菌糸です。菌糸は根を鞘状に覆うので菌鞘と呼びます。その菌糸は細胞間隙に潜って根の中まで入り込むので(写真のHn)、根の細胞は菌糸に抱かれ包まれたような格好になります。その濃密な接触の中で、植物からは、地上部の光合成でできた糖類が菌類に供給され、菌類からは、土壌中に存在する窒素・リン酸・カリウムなどが根に送られます。互いに都合のよい関係にあると理解され、「菌根共生」と呼ばれています。
ところで、子実体が、地中深くから地表までどうやって迷わずに上がってくるのか(写真左)不思議ですね。これは負の重力屈性によります。では、仮に、菌糸が増殖しているところ(排泄所跡)の直下に空間があり、光が真下からやってきたらどうなるでしょうか?子実体は、真っ逆さま、地球内部に向かって生えます。正の光屈性です。これらは、実験室で明らかにされました。このように、菌類はリアリストです。知能(インテリジェンス)があるとも言われています。「リアリストでインテリジェンスもある」という言い方をしてみると、菌類がいっぱしの存在に見えてくるのではないでしょうか。
3. 私のきのこ研究のはじまり-アンモニアとの出会い
農学部4年生の時、アカマツ林にどんなきのこが生えるかという調査をしたのが研究の始まりです。ある時、師匠の濱田稔先生が雑談の中で、「ありのままの自然を調査するのもいいが、山を一つの生物体と考えて、いろいろ刺激を与えてみてはどうか?」と言われました。「面白い」とは思いましたが、具体的にどうすればよいか、すぐにはわかりません。そして、きのこが少しわかるようになってから、山の反応を、きのこを通して観ようと考えました。そして、刺激の方法として土壌への化学薬品施与を考えました。この時、きのこが反応する予感があったわけではなく、文献も見ませんでした。化学薬品としては、肥料を使いましたが、肥やすという考えでなく、安く大量に使えるからでした。そもそもきのこは植物ではありませんから、「窒素・リン酸・カリウム」という肥料の常識は通用しません。ただ、マツタケ増産へのヒントはないか? という考えがほんのすこしありました。
数種類の肥料をアカマツ林の、幅50センチ、長さ10メートルの帯状区画に撒きました。すると尿素施与区に見たこともないきのこが一面に生えたのです(スライド3)。のちに和名を「イバリシメジ」としました。

(スライド3)
尿素を撒いた場所には、いろいろなきのこが長期にわたって生えました。一部を写真でお見せします(スライド4)。AとBはチャワンタケ類、CとDはヒトヨタケ類です。それぞれ種類が沢山ありますが、その中で決まった種類が生えます。新種もありましたし、新種でなくても、尿素を撒いたら生えるというような性質は知られていませんでした。
.jpg)
(スライド4)
初期に生えるきのこは菌根共生をしません。時間が経つと、食欲の対象になりそうな大きなきのこが生えます(スライド5,6)。アカヒダワカフサタケ、ナガエノスギタケダマシなど、どちらも菌根共生をします。
.jpg)
(スライド5)
.jpg)
(スライド6)
ところで、そもそも尿素とはどのような物質で、尿素を撒くときのこが生える真因は何でしょう(スライド7)? 尿素は、皆さんご承知の通り、我々の尿の中に排泄されます。肉や魚などとして食べた窒素分の最終形態です。工業的にはアンモニアと二酸化炭素から合成されます。土に撒くと水に溶け、微生物の尿素分解酵素の作用でアンモニアと二酸化炭素になります。このアンモニアが一番の大元であることがわかったので、尿素を撒いて生えるきのこをまとめて「アンモニア菌」“ammonia fungi”と呼ぶことにしました。世界に約50種類あります。150万種とも言われる菌類全体の中では、一握りにもならない一つのグループです。

(スライド7)
アンモニアが基本であることを示す実験として、アンモニア水を撒くと尿素と同じ効果があります。アンモニアは「土壌殺菌剤」とされたこともある毒物ですから、尿素を撒くということは「殺す」という側面を含んでいます。また、アンモニアはアルカリとして、落葉落枝など有機物の分解を促進します。硫酸アンモニウム(硫安)は「アンモニア性肥料」と言われますが、これは効果がありません。しかし、これを撒いた上から強アルカリを加えると、強酸との結びつきが切れ、アンモニアが有効になります。山林土壌という得体の知れない媒体を通してもなお、化学通りにきのこが動く場面にいくつも出会って感動しました。
4. 動物の排泄物や死体のゆくえ
はじめに戻りますが、イバリシメジの生えた実験(スライド8)で肝心なのは、尿素という物質を撒いただけで、胞子など、菌の種(たね)になるものを撒いてはいないことです。きのこはそこに潜在していたわけです。写真の、白線の手前の非処理地にもイバリシメジは居るということです。

(スライド8)
では、自然界ではイバリシメジはどんなところに生えるでしょうか? 先ずは、尿が落ちたところを調べました。例えば、ヒトの放尿跡にイバリシメジ、イバリスイライカビが生えました(スライド9)。尿素を撒いた時と同様に、落ち葉が黒化しています。

(スライド9)
タンパク質も分解するとアンモニアが生じるのであれば、動物死体でもきのこが生えるだろうと実験しました。安い冷凍の魚を山へ持って行き、腐らせてみるとイバリシメジなどが生えました。豆腐もタンパク質ですから腐れば生えるはずで、それも確かめました。
イバリシメジには特技があります(スライド10)。胞子を寒天培地に撒くと栄養菌糸が伸び、時間が経つと、子実体ができることもあります(写真左)。普通、きのこは傘の下面のひだに胞子をつくりますが、このきのこは栄養菌糸から直に胞子をつくってしまう(写真右)。細い栄養菌糸から太い菌糸が少し伸びてその先に胞子をつくります。この特技は、例えば虫の糞や死体などの微少なアンモニア源からでも胞子までつくれるという性質ではないかと想像しております。
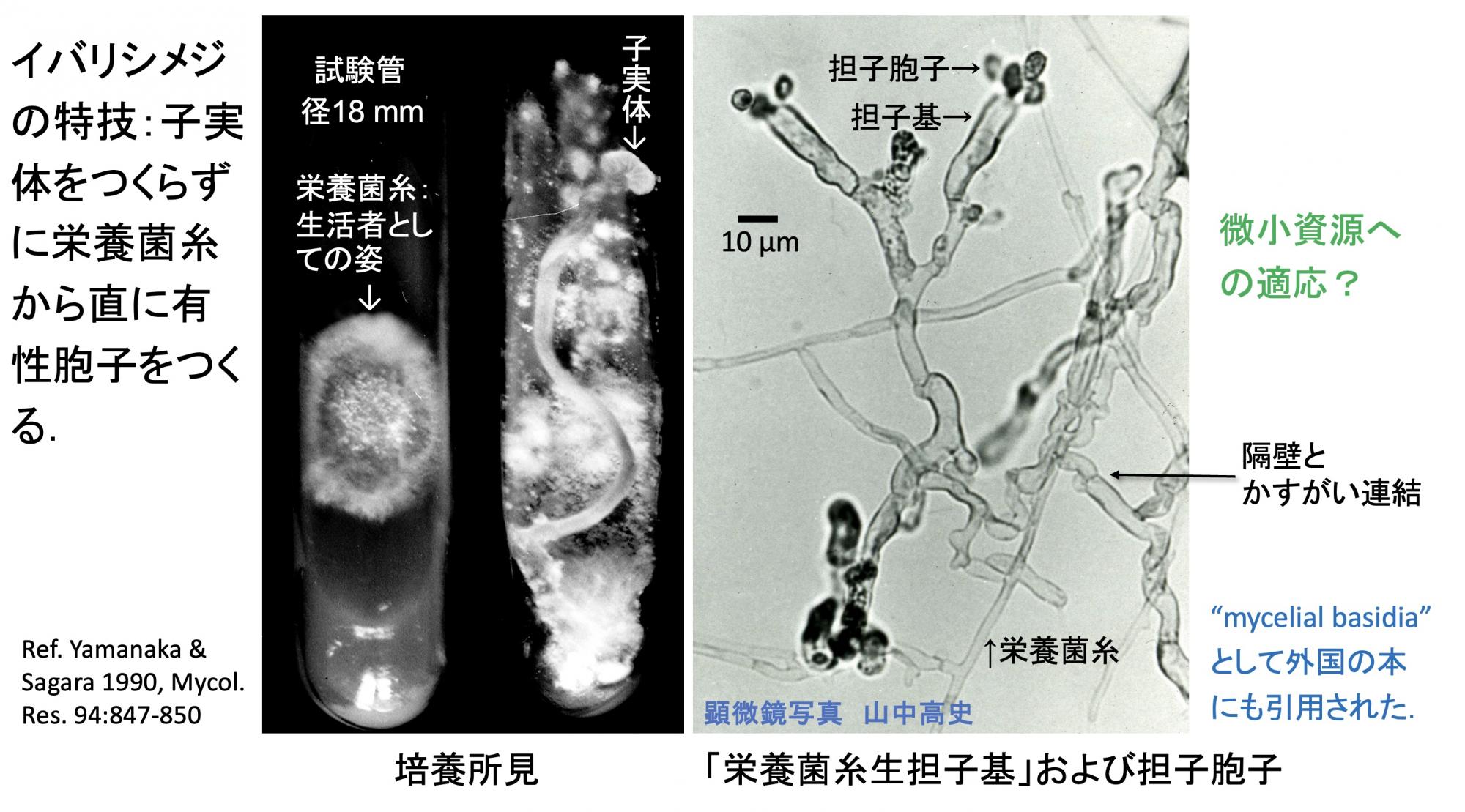
(スライド10)
時間経過に伴って遷移後期に生えてくるきのこの例では、アカヒダワカフサタケが猫の墓に生え(「ごあいさつ」で永田館長が言及した『Nature』はこの知見)、ナガエノスギタケダマシ(当時はナガエノスギタケ)がキャンプ場の野外便所跡に生えました。また、アシナガヌメリがクロスズメバチの巣跡に生えた例もありました。これは、巣の中に溜まったハチ幼虫の糞塊に含まれる尿酸がアンモニア源だとわかりました(スライド11)。
.jpg)
(スライド11)
森の中での動物性廃物の成り行きを模式化して見ると(スライド12)、死体や排泄物が朽ち果てた跡に、きのこの菌糸と木の根が茂って菌根共生し、その菌根の塊がエネルギー源になって地表に子実体として現れます。ここに暮らす動物は、樹木ときのこに生活の後始末をしてもらっているとみることができます。私はそれを「生息地浄化共生」と呼んでいます。
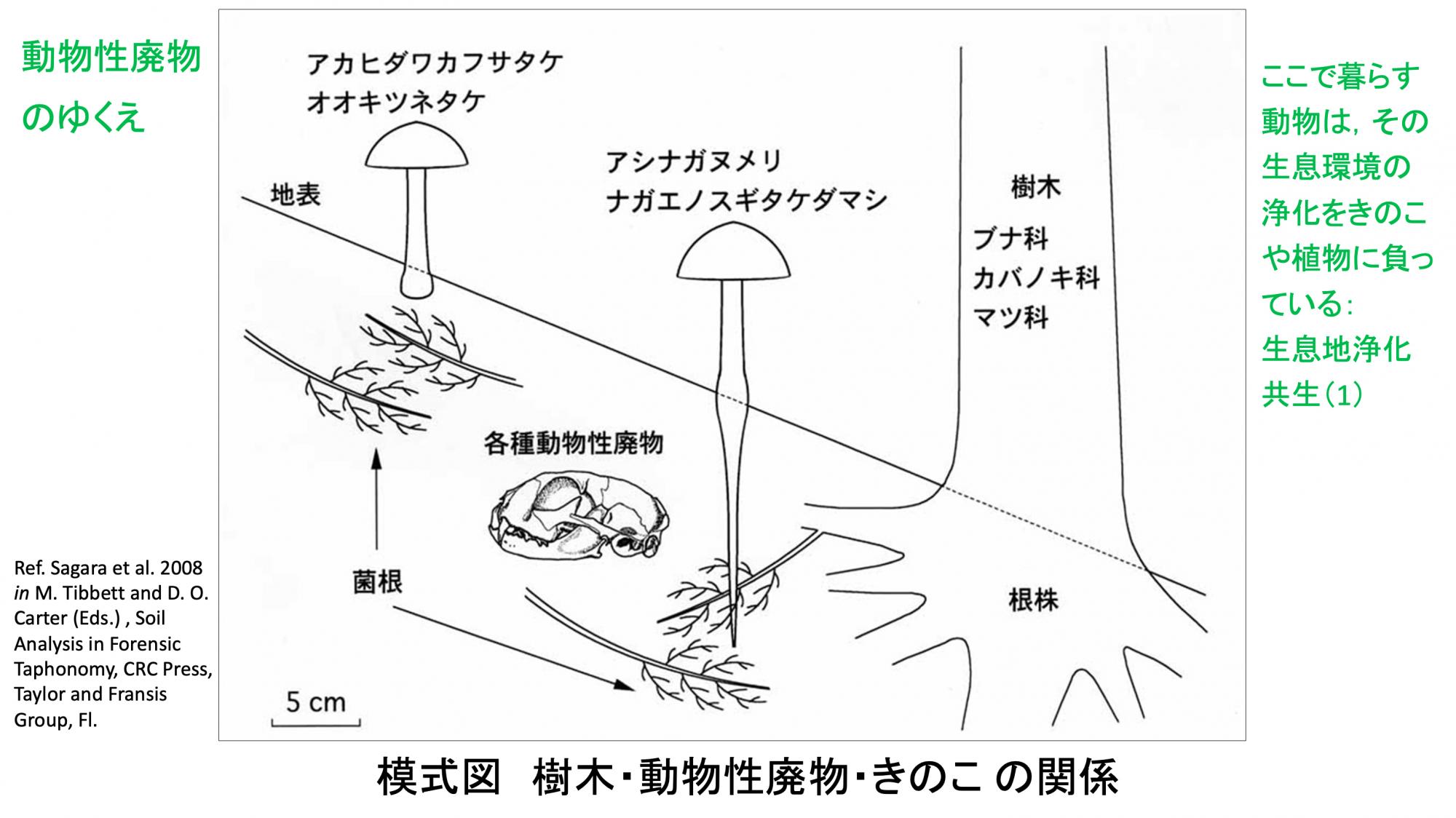
(スライド12)
動物の糞に生える糞生菌は昔からよく知られています。とくに草食動物の糞は分解に時間が掛かるために、長い間、地表に形が残ります。そこに生えるきのこは注目されやすく、たくさん研究されました。アンモニア菌はそれらとは違います。肉食や雑食動物の糞や動物死体はすぐに腐って消えてなくなりますし、尿は初めから形を留めません。アンモニア菌はその跡の土に生えるものです。
5. 2種のきのこの混同-モグラとの出会い
ナガエノスギタケダマシとナガエノスギタケ(スライド13)。今、この写真だけ見ると、どうして両者を混同していたのかと思われるでしょうが、歴史の事実として、混同され、同種だと考えられていました。

(スライド13)
京大近くの大文字山で、本物のナガエノスギタケに出会った時、ナガエノスギタケダマシだと思っていたから、地中にアンモニア源があるはずと考えて堀ったところ、巣に行き当たりました。きのこが生えた理由は動物の排泄物だとすぐに見当がつきました。しかし、巣の主(ぬし)がわからない。動物学者を訪ね歩いたところ、ネズミだろうということでした。動物学者から、「モグラ」の線は全く出ませんでした。当時、「山にモグラがいる」という常識もなかったのです。ネズミの巣と誤同定をして発表してしまいました。その後に、モグラだったことがわかり、ほぼ2年後に訂正しましたが、この大失態のせいもあってモグラに深入りしました。

(スライド14)
二つのきのこを別種ではないかと感じた初めは培養所見です(スライド14)。試験管内の寒天培地でそれぞれの菌糸を培養したところ、ナガエノスギタケダマシは寒天表面にくっついて伸びるから透明感がある。一方ナガエノスギタケは気柱に立ち上がるので白い塊に見える。胞子の形態にも微妙な違いがあることがわかりましたし、さらに子実体の香りもうんと違いました。香気成分を調べると、前者はほとんど持たないのに対して、後者では3物質が顕著に検出されました(スライド15)。うち、上の二つを混ぜ合わせるとナガエノスギタケ特有の香りになることもわかりました。
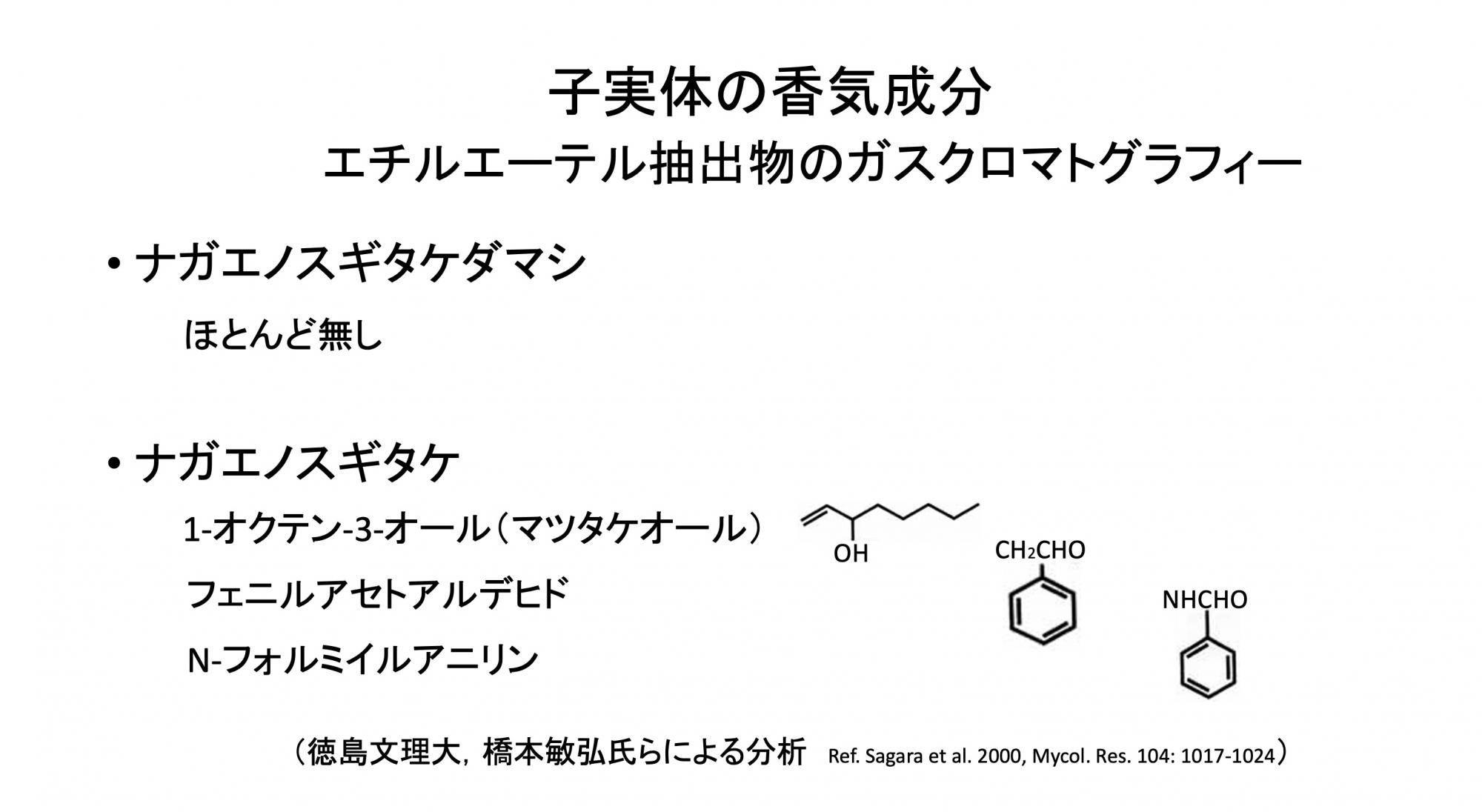
(スライド15)
さらに実験的に、モグラの便所跡に似せて尿素などを地中深く埋めてナガエノスギタケが生えるかと調べてみましたが、そこに生えたのはアンモニア菌のナガエノスギタケダマシとアシナガヌメリのみ。ナガエノスギタケはモグラの便所跡でしか確認できませんでした。このように、騙されていたおかげで私はモグラに出会うこともできたわけです。なお、ナガエノスギタケは室内培養によって子実体を生やすことはできますが、山で人為的に生やすことは今でもできておりません。
そこで、ナガエノスギタケダマシをナガエノスギタケとは異なる新種として発表しました(Sagara et al. 2000, Mycol. Res. 104: 1017-1024)。私が筆頭著者として新種発表を行った唯一の例です。2種の違いに気が付いてから約18年後の2000年のことで、この時、ようやく「ナガエノスギタケダマシ」という和名も定まりました。なお、その後の研究で、ナガエノスギタケとナガエノスギタケダマシは分子系統学的には近縁ではないことがわかっています。
この論文には新種発表とは別の冒険がありました。それは、英語圏外の私がつくったammonia fungi(アンモニア菌)とchemoecological group(化学生態学群)という言葉を、論文のタイトルに使って、定着・普及させようという企みです。審査において何か言われるかと緊張しましたが、何も言われませんでした。「学問は遊びだ!」というのが私の基本姿勢ですが、闘わなければならないこともあるように思います。

6. きのこ - モグラ学
モグラの巣を見つけるには、きのこを手がかりにするほかありません。皆さんご存じの「モグラ塚」は、地下鉄工事の残土を積んだようなもので、あの下に巣があるわけではありません。捕らえたモグラに発信器を取り付けて巣を見つける方法はありますが、科学機器を使わず巣を見つけるにはナガエノスギタケとアシナガヌメリを手がかりにするしかありません。きのこ屋にモグラ学への出番が生じた次第です。

(スライド16)
モグラの雪隠から生えるきのこは、ナガエノスギタケとアシナガヌメリの2種です(スライド16)。アシナガヌメリはアンモニア菌の1種で、いろんなアンモニア源から生えます。ナガエノスギタケはモグラの雪隠専属です。一方のモグラにはこういう種類がいて(スライド17)、どのモグラの便所跡からもナガエノスギタケ、アシナガヌメリは生えます。
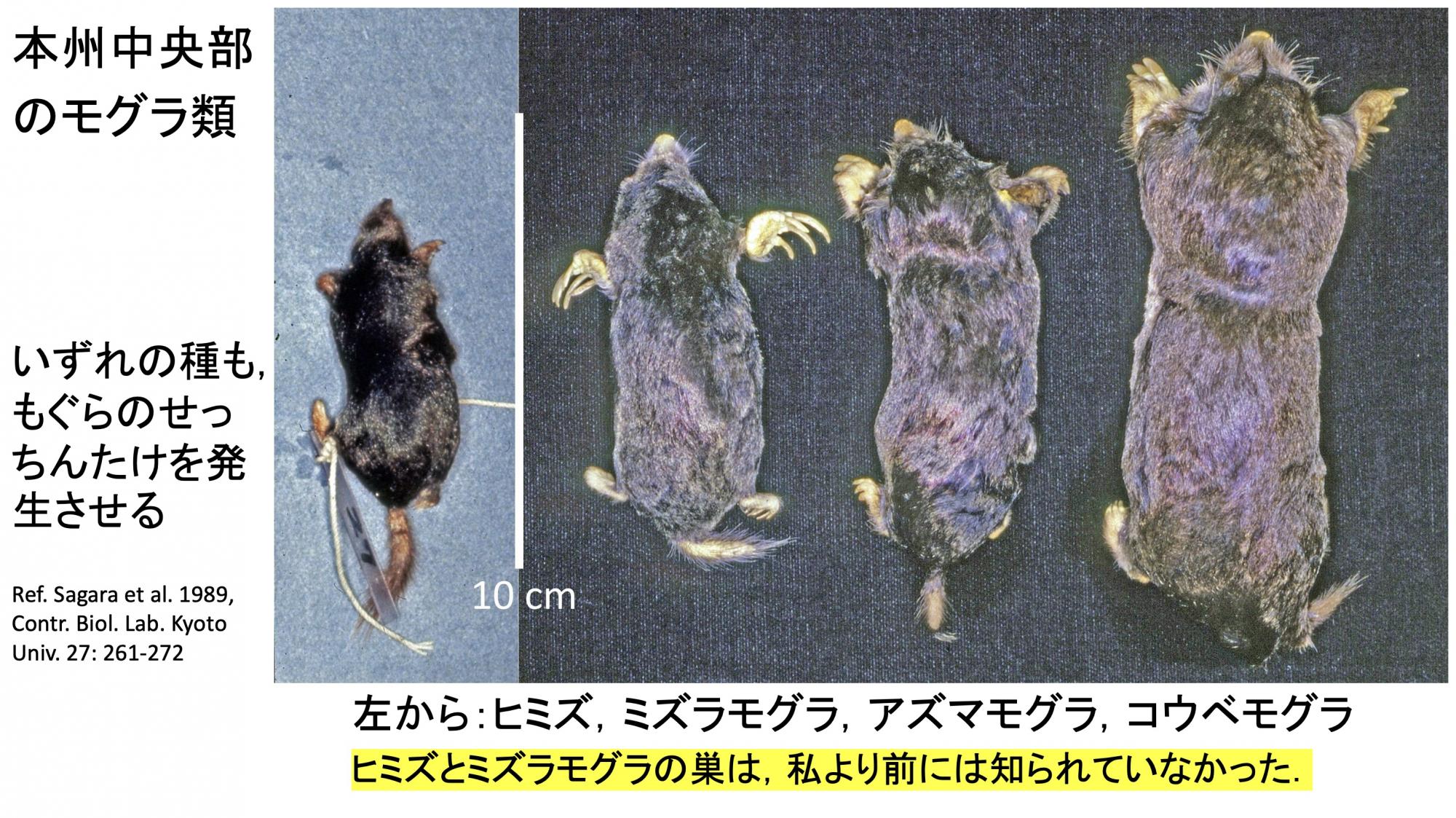
(スライド17)
さて発掘です。これまでに、外国を含めて86箇所、計134回の発掘を行いました。同じ場所でくり返し掘ることもあるので、回数は場所の数より増えます。
この1枚の写真にきのこが生えた由来を示します(スライド18)。画面中央のやや上部の地表にきのこがあります。その下に地中柄があり、便所跡、巣、坑道が示されています。発掘では、このような写真が撮れるように掘ります。この時も、初め、向こう側から掘ったものの、それではよい写真が撮れないとわかってこちら側から掘り直しました。

(スライド18)
なお「モグラが便所をつくる」、もう少し正確に言えば「ほぼ場所を定めて排泄する」という認識は私より前にはありませんでした。排泄物が「臭い付け」に使われるという理解はありました。
排泄物が新しいうちは蛆虫(うじむし)やバクテリア等が増殖し、時間が経つと、きのこの菌糸と木の根が茂って菌根形成をしながら排泄物の後始末の仕上げをします。そこに溜まったエネルギーで地表にきのこの子実体が現れる。ですから、地表で見るきのこはモグラの排泄物の「成れの果て」とみることができます。
ここの諸関係を整理すると次のようになります( 冒頭の図「 樹木・モグラ・きのこ の関係 」を参照 )。
1) 樹木は森を形成してモグラときのこに生息地を提供し、菌根共生によってきのこと養分のやりとりをする。
2) モグラはきのこと樹木に栄養分となる排泄物を与え、土壌の耕耘や通気をもたらす。
3) きのこは樹木と菌根共生を行いながら、モグラの排泄物を清掃する。
森に棲むモグラは、このように、樹木やきのことの共存から恩恵を受けていると考えられるので、この関係も「生息地浄化共生」と捉えます。特に長期の定住において、このことは重要ではないかと思います。
巣は、モグラの日々の暮らしの中心であるにもかかわらず、これまで真面目に観察されませんでした。ヨーロッパの本を見ると、巣は、出入口が二つあり、落ち葉の雑然とした塊に過ぎず、モグラはその中に体を放り込んで休息するという程度の理解しかありませんでした。そうではなく、巣の出入り口は一つであり、巣には決まった構造があることを、私は明らかにしました(スライド19)。巣は、広葉樹の落ち葉を地中空間の壁に押し当てて、しっかりとつくられています。スライドの写真(上)は、病院の、ヒトをのせるベッドにモグラの巣をのせて撮影してもらったものです。ここで見られるような巣のつくりに、モグラにおける「文化」を見る思いです。
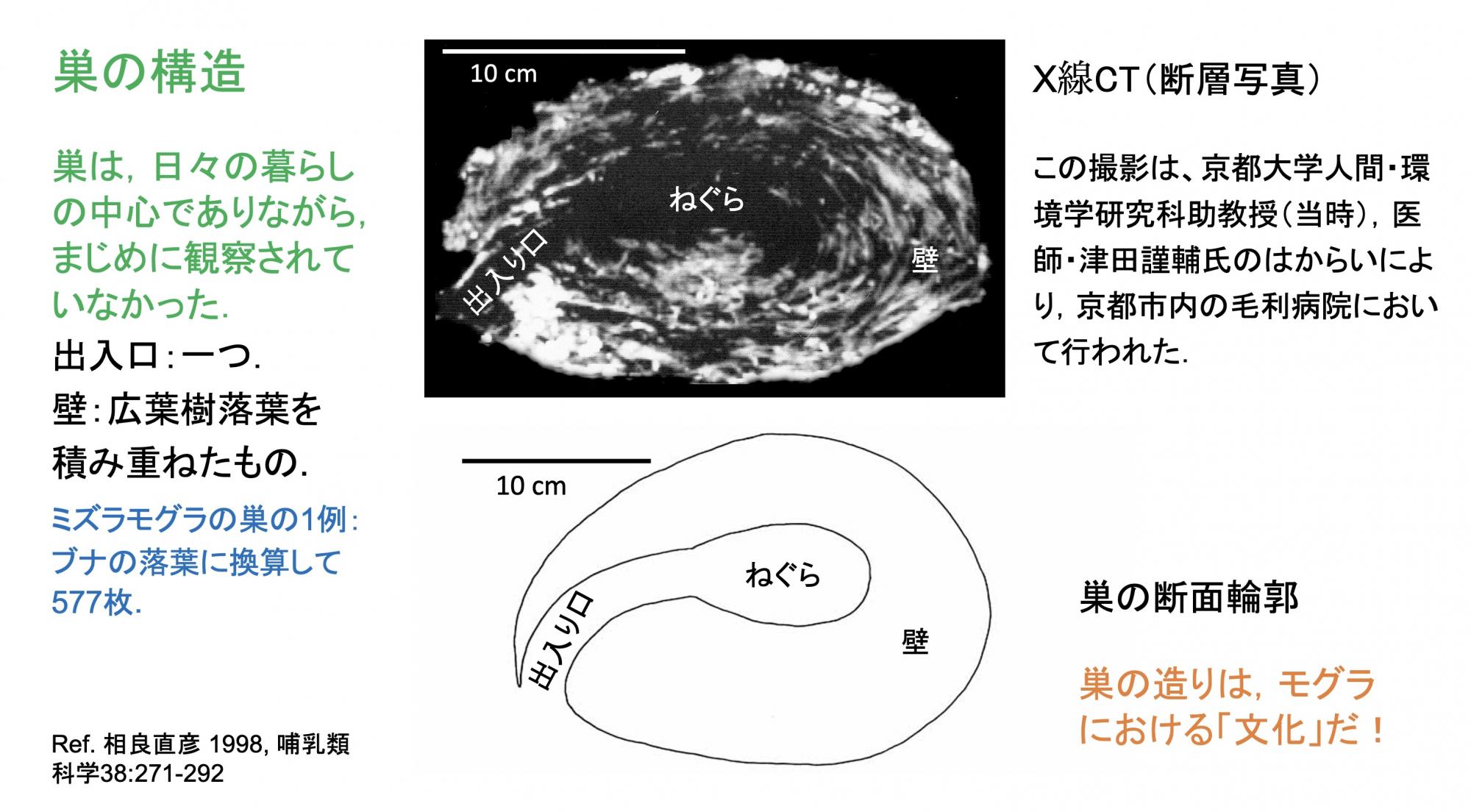
(スライド19)
私のモグラ学でもっとも大事なところは、モグラは長期に定住する傾向があることを明らかにしたことだと思っています。モグラの寿命は数年とされるなか、20年以上にわたって同じ場所で営巣がおこなわれた例があります。その話をここで詳しくする時間はありませんが、住者が代わってもなお同じ場所に住み続けた証拠をひとつお話しします。
京大の芦生研究林で、1985年にナガエノスギタケが生え、その翌年に発掘し、巣とその住者をともに採取して埋め戻しました。住者はコウベモグラでした。そこから9年後の1994年、同じ場所にまたナガエノスギタケが生えました。巣も住者もいったん取り去っていますから、別のモグラが営巣したに違いありません。掘ってみると、巣があり、今度はアズマモグラが住んでいました。モグラの種類が変わってもなお、同じ場所で営巣することがあるとわかりました。異種のモグラが近接して住んでいる証拠でもあります。ところで、「長期定住」と言う時、私が見た期間が20年あまりであって、その前、いつから営巣がはじまったのかはわかりません。

(スライド20)
長期定住の観察の中で、予め場所がわかっていたミズラモグラの巣を掘ったところ(スライド20)、幼獣がいました(写真左上)。その時は、写真を撮っただけで埋め戻し、後日、再び掘ったところ幼獣はこのように成長していました(写真右上)。このうち一匹を家(京都)に連れ帰り、餌付け・飼育を試みました(写真左下)。まだ乳離れしておらず歯も未発達で、ミミズを丸ごと与えても食べられない。ミミズをはさみで切って食べさせたりして飼育を確立して(写真右下)、その後は、東京の本職の哺乳類学者が飼育・観察を継続しました。そして最後は、きれいな本剥製標本として返却されました。その標本は、今は京大博物館に展示されているそうです。
モグラは「農耕地周辺の動物」と思われがちですが、農耕地が開かれたのは地球史上では近年のことであり、見てきたように森にも普通に住んでいて樹木やきのこと深い関係を結んでいますし、広葉樹の落葉を材料としてつくられる巣も完成度が高いと感じられるので、「モグラは、本来、森の生物だ」と私は主張しています。
7. ナガエノスギタケを追ってヨーロッパへ
ナガエノスギタケはモグラの便所跡から生えることが日本でわかりましたが、イギリスでも同じ例を私が明らかにしました。一方、モグラがいないのにナガエノスギタケが記録された地域がありました:アイルランド、ノルウェー、それから日本の北海道です。それらの地域で、ナガエノスギタケが生えた原因が問題になります。調査の結果、北海道ではトガリネズミの例が見つかりましたが、トガリネズミは広い意味のモグラ類ですから、これはあまり不思議ではありません。一方、スイスではモグラとネズミそれぞれの例が、またノルウェーではアナグマの例が見つかりました。ネズミの例は意外感があります。食性が大きく違うからです。なお、アイルランドは調査できておりません。
ところで、こうした調査が可能だったのは、きのこ発生地点をピンポイントで標識し、連絡し、現地案内してくれる協力者が存在したからです。日本国内でも同様で、私が調査した73箇所のうち、自分で見つけたのはわずかに12箇所で、他はきのこ仲間からの通報によるものでした。ここではスイスからの通報に基づくモグラとネズミの例を紹介します。

(スライド21)
モグラの例です(スライド21)。きのこ発生地点は、概観では「針葉樹林」の中でした。「針葉樹のマツ科もナガエノスギタケと菌根共生を行うのだろうか?」という問いを抱えていましたが、現地に着いてみるとナガエノスギタケが生えた付近には、落ち葉を散り敷くブナの木が3本ありました。トウヒの根元を掘って巣を掘り出し、中に落ちている毛を調べたところ住者はモグラであることがわかりました。巣材はブナの落葉でした。このように概観は針葉樹林でも、ナガエノスギタケが生えた場所には必ずブナが存在したという例を4箇所経験し、ブナ・モグラ・ナガエノスギタケという三者の結びつきの鮮明さに感動しました。なお、「マツ科はナガエノスギタケの菌根宿主になりうるか?」という問題が解消したわけではありません。発掘作業は、私の “Site-seeing”であり、大地の秘密と向き合う至福の時です。この時の気温は1℃でした。

(スライド22)
もう一つは、ブナ林でのネズミ(ヨーロッパアカネズミ類)の例です(スライド22)。私に連絡をくれた、赤い上着の人は菌学者で、彼女の知り合いのきのこ愛好家(写真左)がしょっちゅう山を歩いていて、ナガエノスギタケを見つけて標識しておいてくれたわけです。標識の下を掘ってみると、巣は割合浅いところにあり、そこから坑道でつながった深い所に、ブナの実の食べかすなどが大量に溜まった貯食痕がありました。モグラの場合、こういうものはありません。住者は、やはり毛を調べることでアカネズミ類だとわかりました(写真右)。調査の後、愛好家のファールニさんは、「たった1種類のきのこのために、遠路はるばる日本から、本当にやって来るとは思わなかった」と言いました。
8. 結び
結び、1.きのこは過去を語るということは、十分ご理解いただけたと思います。「きのこには証拠力がある」とも言えるかもしれません。そこから鑑識菌学・法菌学というような分野への広がりも予感されます。分野の確立には至りませんが、「証拠力」に力点を置いて、尿素施与に始まる研究をまとめました:N. Sagara, T. Yamanaka and M. Tibbett. 2008. Soil Fungi Associated with Graves and Latrines: Toward a Forensic Mycology. In: M. Tibbett and D. O. Carter (Eds.), Soil Analysis in Forensic Taphonomy, CRC Press, Boca Raton, FL. この一篇は、米国で出版された本の中の1章で、その本を企画したイギリス人研究者の誘いで書きました。私(たち)が書いたものの中では一番よく読まれているようです。
結び、2.献名いただいたきのこの学名が3つあります。Panaeolina sagarae Hongo (コブミノシバフタケ)、Sagaranella V. Hofstetter, Clémençon, Moncalvo & Redhead (イバリシメジ属)、およびHebeloma sagarae T. Kasuya, Mikami, Beker & U. Eberh. (ナガエノスギタケ) です。ナガエノスギタケは近年になって、ヨーロッパのものとは違うという見解に至り、日本のものは新種で、新しい種小名が必要となったわけです。
献名は、虫にも一例ありました。Lucasioides sagarai Nunomura (モグラアナワラジムシ)。これは、富山県で私も関わって掘ったモグラの巣の試料から得られたものです。きのこからモグラの巣を見つける道が開けたことからこんなものまで生まれたのは、とてもhappyです。

(スライド23)
結び、3.これは何をやっているところかおわかりでしょうか?(スライド23)。この桶(おけ)は肥担桶(こえたご)と言いまして、下肥(しもごえ)、つまり我々のし尿を汲んで畑に入れます。昔はどこでもやっていたことです。この作業は、「一滴も溢さず、一抹も浴びないぞ!」という集中力が要る作業でして、これをやり終えると、人としての勤めを果たしたような思いで気分爽快になります。
― 謝辞 ―
きのこ発生情報をくださった方々をはじめ、研究活動の中でお世話になった方々、研究資金を援助くださった富士電機(株)様に厚くお礼申し上げます。その方々のおかげで成り立った「私のきのこ学」でありました。田舎ごもりの私にこの講演の機会をくださった永田館長、および準備を助けてくださったスタッフの方々にお礼申し上げます。そして会場の皆さん、それからインターネットで聴講くださった皆さん、ありがとうございました。最後に、骨肉となった郷里とその人々、および犠牲になりながら助けてもくれた家族への謝意も表しておきたいと思います。

相良直彦
(さがら・なおひこ)
1938年、大分県生まれ。1960年、京都大学農学部卒業。1962年、京都大学大学院農学研究科修士課程修了。1966年、京都大学大学院農学研究科博士課程退学、京都大学教養部助手、助教授、教授、同大学大学院人間・環境学研究科教授を経て、2001年、定年退職。農学博士、京都大学名誉教授。現在、郷里大分県の山間地で独居・百姓。主著に『きのこと動物』(築地書館)。


私のきのこ学
「後始末」を軸としてみる森の姿大分県在住





.jpg)
.jpg)











