顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
神経記憶III
2016年12月15日
神経科学の歴史について詳しいわけではないが、記憶を始めすべての脳機能が神経細胞(ニューロン)同士の結合した回路により担われていることを最初に示唆したのはスペインの生んだ巨人ラモン・カハールだろう。前回述べたように(前回図5)、カハールはゴルジ染色法を用いて、一個一個の神経細胞を染め出し、脳が様々な神経細胞のネットワークであることを明らかにし、カハールの4原則を提案した。このことから、カハールを神経細胞ドクトリン(ニューロン説)の最初の提唱者と呼んでいいだろう。実際この4原則に適合するネットワークを形成できる細胞は神経細胞以外にない。
カハールのニューロン4原則に加えて、神経回路を考える時に重要なドクトリンをあと二つ挙げておこう。
一つはすでに前回紹介したが、神経細胞は共通のメカニズムで、細胞膜上で起こる電気的興奮を伝搬することができる点だ。静止状態では、神経細胞内外のナトリウムイオン(Na+)勾配によって、神経細胞内外の電位差(静止膜電位)は-90mVに保たれている。外的な刺激でこの電位が変化してある閾値を超えると、イオンチャネル(電位に応じて開閉するNaチャネル)が急速に開き、結果急速にNa+が細胞内に流れ込み、膜電位の極性が変わる(+20mV:脱分極)。この電位の変化は、イオンチャンネルを介してK+を細胞外へ逃すことですぐに元に戻るが、刺激が繰り返されるとイオン勾配が消失して、興奮できなくなる(不応期)。このイオン勾配を復元するためには、それぞれのイオンに特異的なポンプが働く必要がある。このイオンドクトリンに従う細胞であれば、いかに複雑な形態をもち、神経間の興奮伝達に様々な化学物質が使われる場合も、神経を点から点に張り巡らせた電線と同じように単純化して考えることができるようになった。
最後のドクトリンが、神経細胞間の興奮伝達がシナプス接合部での化学物質の伝達によって誘導されるとするシナプスドクトリンだ。すなわち、化学物質の伝達によって、それを受け取る側の神経細胞の興奮を誘導できるが、この場合は電位依存性チャネル(voltage gated channel)ではなく、シナプスで興奮した細胞から分泌される伝達物質と結合することで開くチャネルにより、もう片方の細胞に興奮が伝わることがわかっている。シナプスドクトリンは、神経興奮伝達が一方向にのみ進むというカハールの原理を説明するだけでなく、使われる化学物質を変化させることで、神経間の結合に特異性を与え、また次の神経の興奮に対して促進的にも抑制的にも働けることが説明できる。この二つの性質を合わせると、ほとんど無限の神経回路を作れることが直感的にわかると思う。
この3つのドクトリンを基盤にして、神経回路による記憶維持の細胞学的、生化学的メカニズムについての研究が始まったが、この問題を分子レベルまで掘り下げて説明するのに成功したのがコロンビア大学のエリック・カンデルで、彼はこの研究によりノーベル医学生理学賞に輝いている。
カンデルが記憶回路の解明に用いた動物はアメフラシ(図1)で、大きな殻のないカタツムリと考えて貰えばいいだろう。

カンデルは、図2に示した、アメフラシの水管(水を吐き出す管)を刺激すると、エラを引っ込めるという反射に注目した。図2は2000年カンデルがノーベル賞を受賞した時のプレスリリースに掲載されたアメフラシの図だが、腹側にエラと水管が存在している様子がわかる。カンデルたちは、エラを引っ込める反応をアウトプット、水管を触れる刺激、および尻尾への強い電気刺激をインプットとして、それぞれの刺激間の関係や、刺激のパターンが記憶されるかについて調べた。
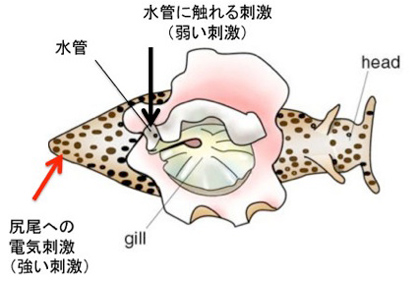
図2:カンデルの用いた実験系。(C) The Nobel Assembly at Karolinska Institute
例えば水管を刺激し続けると、アメフラシは刺激に慣れてエラ反射を起こさなくなる(Habituation)。あるいは、水管に触れる前に尻尾に強い刺激を与えると、水管を触れた時のエラの反応が増幅する(Sensitization)。このサイフォン刺激や尻尾への強い刺激が記憶されているかどうかは、一定のインターバルをおいて、同じ刺激を加え、エラが同じ反応を示すかどうか調べることで確認出来る。このインターバルが短い時の記憶が短期記憶で、長い時の記憶が長期記憶になる。カンデルたちは特にSensitization(感作)と呼ばれる行動を用いて短期記憶、長期記憶について研究を行った。
もちろん短期記憶、長期記憶が存在することは、様々な神経システムで観察することができる。しかし、記憶に関わる神経回路を、一個一個の細胞レベルまで分離することは簡単でない。しかしアメフラシの回路では、神経が大きく、また数が少ないため、エラを引っ込める行動とともに、それに関わる単一の神経細胞の興奮を長期間記録し続けることができる。また、刺激後何日もたって、再度同じ神経を探し出して記録することも可能だ。記憶のメカニズムを、行動、神経生理学、そして細胞内での生化学的反応へと還元したいと考えていたカンデルにとっては、アメフラシは選びに選んだモデル動物だった。
図3はカンデルたちが分離した、水管反射の記憶に関わる神経回路の模式図だ。水管への刺激を感じる感覚神経と、エラを引っ込める運動神経がシナプス結合を形成しており、これがメイン回路になる。そこに、水管の刺激をフィードバックしたり、フィードフォワードする刺激性介在ニューロン、抑制性介在ニューロンが結合してサーキットが形成される。このサーキットに、尻尾の感覚神経と結合する調節介在ニューロンが結合して、体の他の部分の感覚を伝えるまさに回路が形成されている。
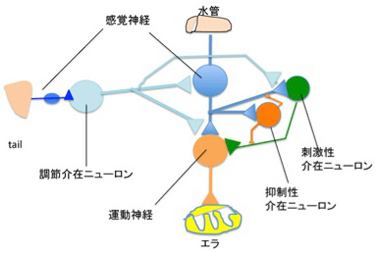
図3 カンデルたちが研究したエラを引っ込める行動を調節する神経回路。実際には、他にも調節介在ニューロンが関わっていることがわかっているが、ここでは省略する。基本的に、水管の刺激を感じる感覚神経と、エラを動かす運動神経の結合した経路に、抑制性および刺激性介在ニューロンが結合し、また全く異なる組織(tail)からの刺激が調節介在ニューロンを介して、この回路に伝えられる。
それぞれの神経は、単一細胞レベルで興奮を記録するとともに、物質を注入することができる。これにより、行動を神経生理学、そして最終的に神経内部での分子間相互作用へと還元することが可能になった。
カンデルたちの期待通り、現在ではSensitizationについての短期記憶、長期記憶のメカニズムはほぼ明らかになったと言える。両記憶のメカニズムについて、同じくノーベル財団のプレスリリースで使われた図を用いて説明しよう。
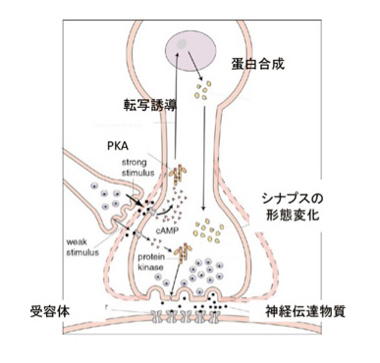
図4:短期記憶と長期記憶。(C) The Nobel Assembly at Karolinska Institute:説明は本文
結論的には、「短期記憶も、長期記憶も、アウトプットとしてシナプス伝達の強さや閾値を変化させる細胞内の変化」とまとめることができる。
まず短期記憶から見てみよう。尻尾からの調節性介在ニューロンは、水管の刺激を感じる感覚神経と運動神経とを結合しているシナプスに結合している(図3)。この介在ニューロンが興奮すると、1)セロトニンを分泌し、2)このセロトニンがシナプス上の受容体に結合すると、3)アデニルシクラーゼが活性化し、4)cAMPが作られ、5)cAMPにより細胞内のタンパク質リン酸化酵素PKAが活性化され、6)カリウムチャンネルや様々なシグナル分子をリン酸化して、7)より多くの神経伝達物質を分泌するシナプスに変化する、という順序で運動神経への刺激が増強する。ただ、この増強は一過的なPKAリン酸化活性の増強によるもので、時間が経てば元に戻る。
これに対して長期記憶は、同じように活性化されたPKAが一過的にシナプス機能に関わる様々な分子を活性化するだけでなく、図4にあるように転写因子を活性化し、誘導された新しい分子により細胞の分化が誘導されることで、長期にわたるシナプスの大きさ数の変化が起こることで維持されることがわかった。実際には、セロトニン刺激がPKAを活性化するところまでは短期記憶と同じだが、長期記憶の場合はその後CREB1と呼ばれる転写因子が活性化され、新たな遺伝子を誘導することで、細胞自体が分化し、細胞及び組織レベルの構造変化が起こることで長く続く記憶が成立する。言い換えると、細胞をプログラムし直し、違った性質に変化させることが長期記憶のメカニズムだ。この仮説は、感覚神経へCREB1を過剰発現させることで、刺激なしでも長期記憶が成立するという実験による証明された。
実際にはこの単純な回路では、さらに複雑な短期、長期の細胞変化が起こり、シグナルのインプットとアウトプットの関係をさらに複雑にできることがすでに明らかにされているが、その詳細は省いていいだろう。
アメフラシの神経回路を用いた短期、長期記憶については以下のようにまとめることができる。
- 1)記憶は、複数の神経がシナプスで結合した神経回路を基盤に成立する。
- 2)シナプスから分泌される神経伝達因子は、細胞の興奮だけでなく、細胞内のシグナル伝達経路を刺激する。
- 3)このシグナルにより、一過性に細胞内の様々な分子が活性化、あるいは不活化され、その結果シナプス興奮の閾値や大きさが変化し、刺激に合わせた反応を誘導できる。これが短期記憶。
- 4)同じシグナルは、特定の転写因子を活性化し、新しい遺伝子発現を誘導する。これにより、細胞のプログラムが変化し、細胞分化や構造変化が起こる。一旦プログラムが書き換えられると容易に元に戻らないため、長期にシナプスの活性を変化させることができる。これが長期記憶。
以上のことから、長期記憶も、短期記憶も、特に特別なメカニズムを使っているわけではなく、多くの細胞と共通の分子を利用していることがわかる。PKAは多くの細胞でcAMP濃度上昇を感知し、短期・長期の細胞変化を誘導するのに働いている。CREB1についても同じだ。ただ神経の場合、刺激により誘導される最終結果が神経細胞同士の神経伝達の強度や閾値の変化になる。またこの変化も、一過性から、様々な長さの時間持続させることが可能だ。
次に、図3からわかるように、神経同士の結合はほぼ無限に複雑化することができる。しかも、神経細胞は形態的にも機能的にも何十種類も存在するため、回路の複雑性はすぐ天文学的になりうる。そして、神経伝達物質や受容体も数十種類存在するため、これによって可能なシナプスを介する興奮伝達の様式も無限に変化させることが可能になる。
しかしネットワークがどんなに複雑でも、背景にある生化学的原理はほぼ共通であることが重要で、どんな複雑な神経系回路でも、刺激が誘導するのは、回路自体の特性のリプログラムだ。
これまで、私の知識を再整理する意味で、神経生物学にあまりに深入りしたが、勉強はこのぐらいにして、次回は神経回路による情報と記憶の特性について考えてみたい。