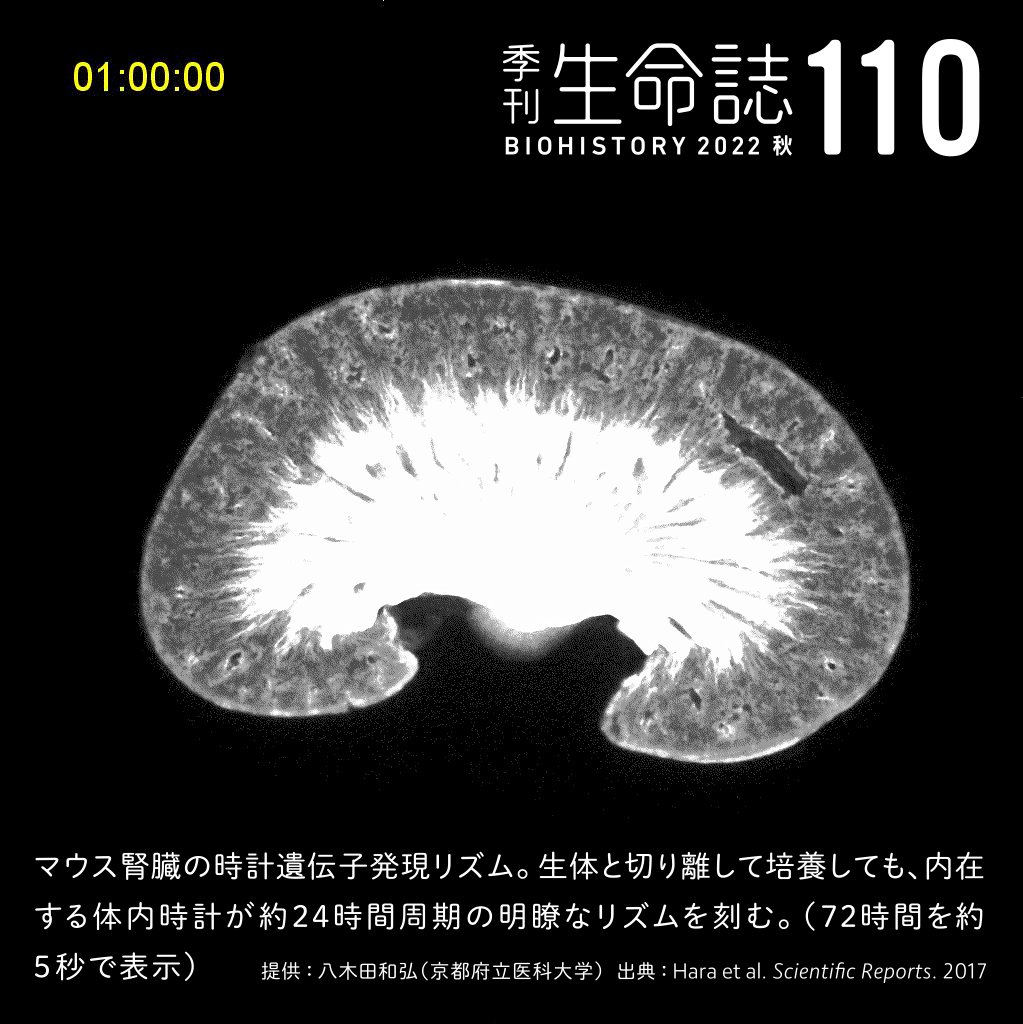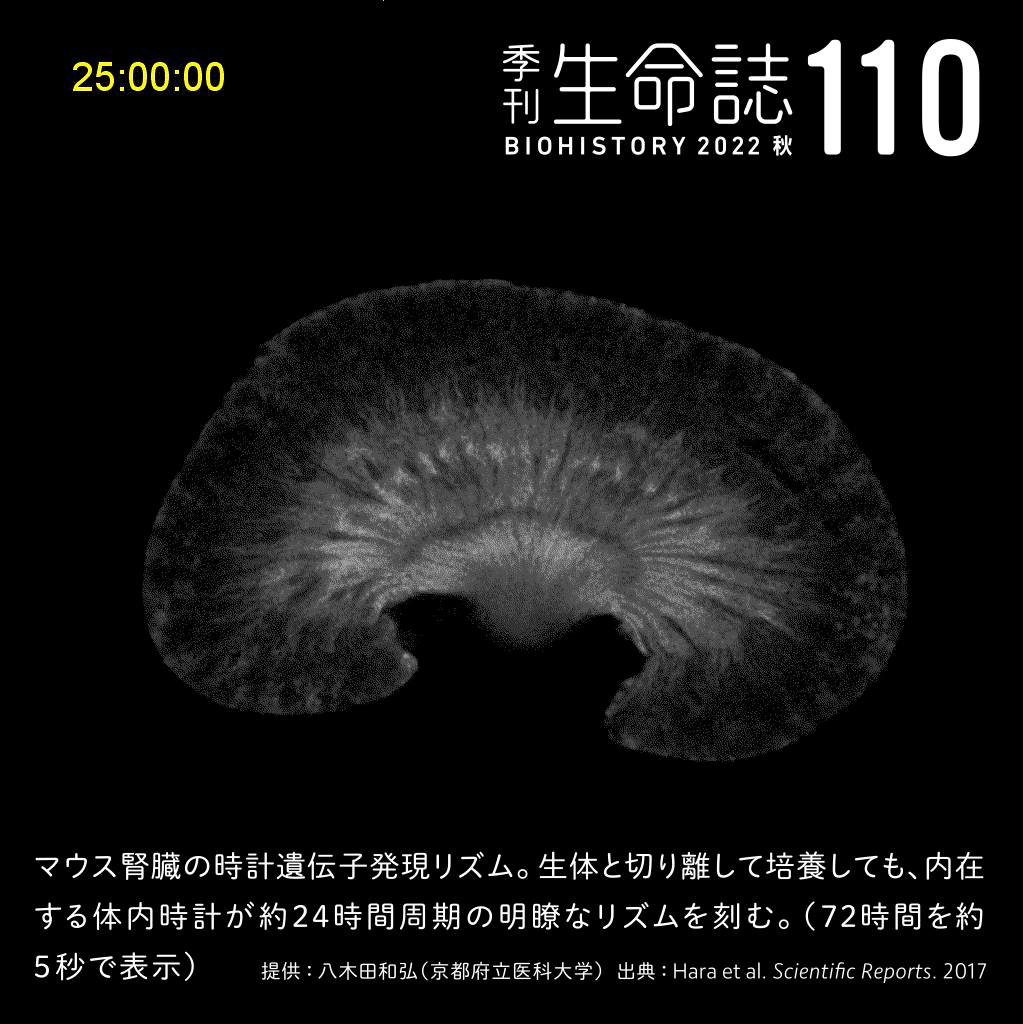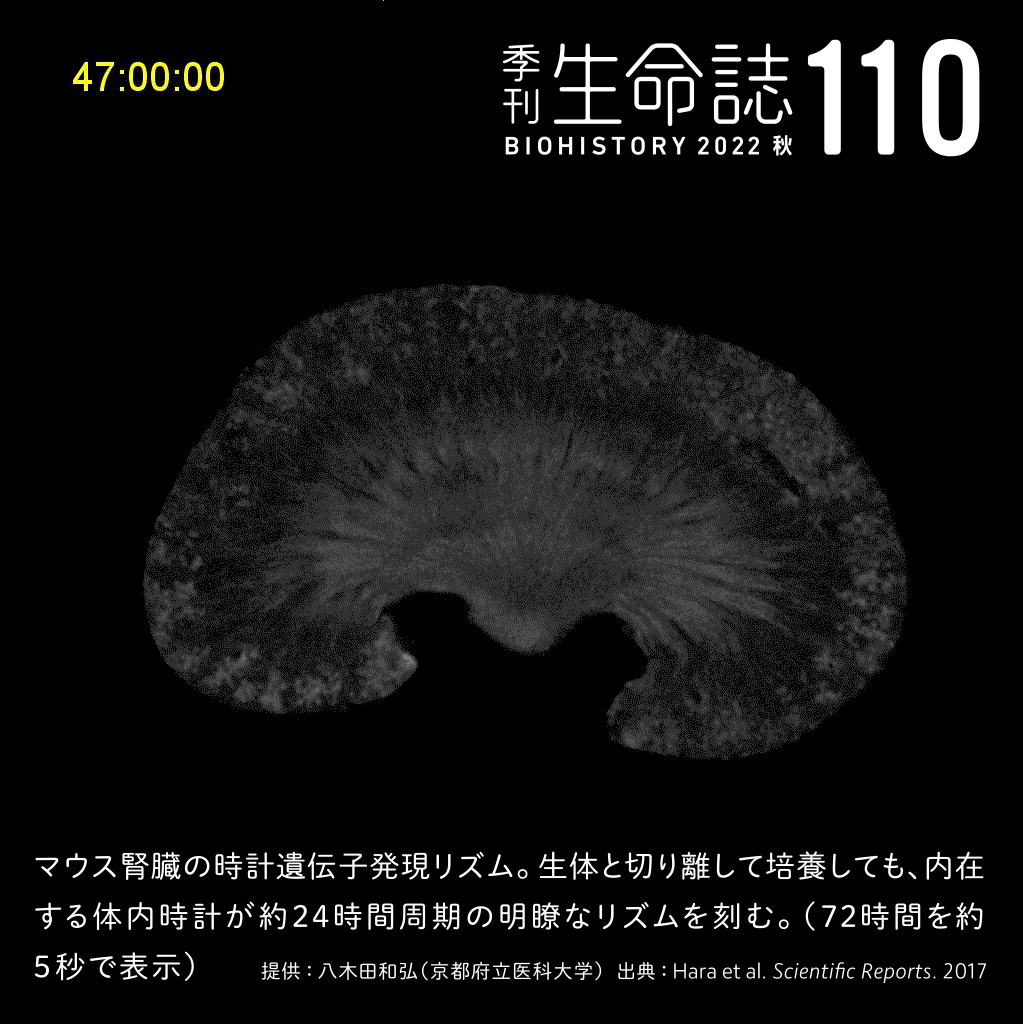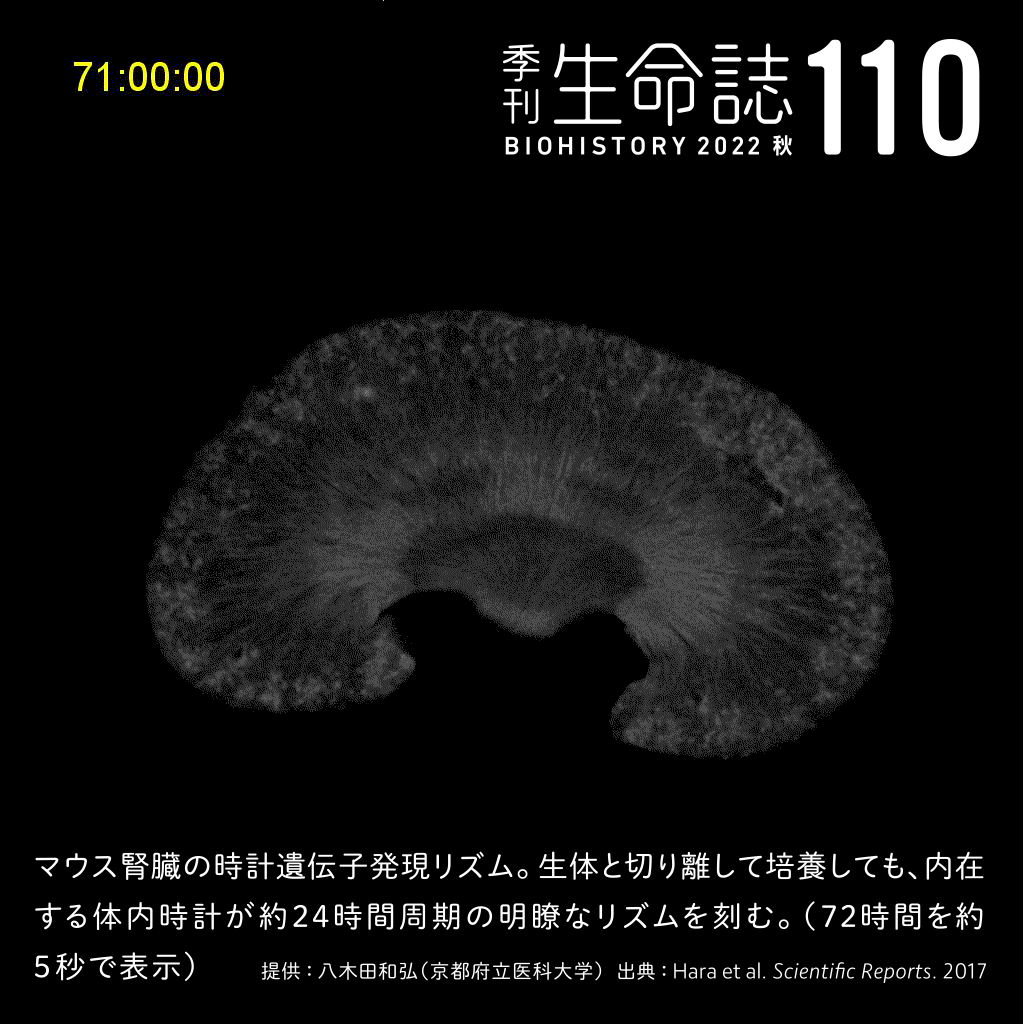
Special Story
オサムシから進化を語る
「バッハは小川(バッハ)ではなく、大海(メール)である」とベートーベンは言い、「バッハは終局である」とシュバイツァーは書いた。しかしバッハは、流れを集めて再び注ぎ出す大きな湖なのだと思う。生命誌研究館で行ったオサムシの共同研究もこれに似ている。個性も歴史も異なる個人(小川)が、1つの目的のために結集し、個人レベルではなし得ない研究(湖)を完成させ、それを糧に再び独自の河川となって流れ出る。どんな湖ができたか、なぜオサムシで、なぜ分子系統なのかを語ろう。
分子系統進化学
名古屋大学在職中、いろいろな細菌でリボソームのタンパク質組成が異なることに気付き、これを利用して細菌の系統を調べようと考えた(1967年)。木村資生(もとお)の中立説(1968年)や、ZuckerkandlとPauling(1965年)のヘモグロビンによる系統樹の話も知らず、分子進化の研究も少なかった時代だ。大学院生の堀寛(現名古屋大学)の協力を得て、大腸菌などの腸内細菌を分析してみると、これまでの細菌学で近いとされていたものは近縁、遠いとされていたものは遠縁となり、なかなかもっともらしい系統樹が書けた(1978年)。これで、分子で系統を追えるという確かな感触を持ち、堀が中心となって5SリボソームRNAを使って約10年がかりで全生物界を包含する分子系統樹を作った(1987年)。DNA塩基配列を決定する方法はまだなく、技術的に難しいRNAで解析せねばならなかったが、古細菌が真核生物に近いということを初めて指摘することができ(1979年)、新たに後生細菌という名を提唱した。また、分子を使って系統進化を研究する新しい分野を「分子系統進化学」と名づけた(『自然』1980年5月号)。
オサムシでいこう
退官近かった1991年のある日、私は国立岡崎共同利用研究機構長の岡田節人(ときんど)(現JT生命誌研究館館長)と名古屋のホテルで会っていた。
岡田
近々、中村桂子(現同副館長)と一緒に生命誌研究館というユニークなものを作るんや。あんたも名大をやめたら参加してくれんか
大澤
何をやればいいんですか
岡田
何もせんで遊んでいてくれたらええんや
幸い、虎の門の生命誌研究館準備室では昆虫を研究対象にするつもりで、オサムシの形態分類の本をまとめたばかりの石川良輔(当時東京都立大学)から話を聞いているということだった。少年の頃から愛してきた甲虫で研究ができるなら、まさに遊び心だ。そこで分子系統の研究をしようということになり、最初はテントウムシも候補に考えた。しかし、これは種類が多すぎる。結局オサムシに落ち着いた。その時点で私は、形態や分布の知見から、オサムシ多様化のストーリーを頭に描いたが、その後のDNA解析の結果はそれとは大きく違い、客観的データに基づく実証の重要性を 痛感している。
やはり昆虫少年だった岡田は、 「生命誌研究館は一般の人にも楽しんでもらうことを狙っているので、北海道のオオルリやアイヌキンなどの派手なのを是非忘れんように」という館長としてのコメントも忘れなかった。
開館までの間、名大で乾燥標本のDNA解析を試したが、全て失敗。そんな折、哺乳類の寄生虫のDNA解析のことで研究室を訪れた岡本宗裕(当時大阪大学)が、オサムシの予備実験の話に目を輝かせた。ムシ屋に急変した岡本は、北海道大学時代、オオルリオサムシやアイヌキンオサムシをたくさん採っていて産地にも詳しいと言う。かくて、岡本はオサムシプロジェクトの共同研究者第1号となり、岡田所望の派手なオサムシの解析のめどがついた。

オオオサムシ(写真:大西成明)
生命誌研究館で
93年、生命誌研究館開館。系統樹作成に、最初は核の遺伝子を使うつもりでいたが、進化速度が遅く、オサムシのような小グループには不向きとわかり、ミトコンドリア(mt)DNAの中で、進化速度が速いとされるND5遺伝子を選んだ。乾燥標本は使えないので、アルコール漬けのオサムシで試すと100%成功。石川の試料で予備的な分析をし、1年かけて技術的な問題を解決した。
94年、DNAの扱いに精通した大濱武(1年後オサムシから離れ共生進化の研究に専念)と、蘇智慧(スーズィフィ)の加入で、分析は急速に進み、半年を経ずしてかなりの結果が出た。しかし、その結果たるやまったく予想外で、これまでの形態分類の常識とかけ離れている。石川に試料をもらいにくくなったので、名大学生時代の虫の先生だった故中根猛彦に窮状を訴えると、「生きた材料集めにはアマの研究者や研究機関への依頼が早道」と、大阪市自然史博物館の宮武頼夫館長を紹介された。日ならずして、宮武、冨永修、桂孝次郎ら近畿オサムシグループが生命誌研究館へ来訪、協力を約束してくれた。
日本のオサムシの生き字引である冨永は、オサムシの産地を熟知しており、必要な試料を次々と届けてくれた。生命誌研究館での解析結果を伝えると、的確なコメントと共に、不足の試料が調達されて返ってくるのだ。このキャッチボールは今なお続いている。冨永といい、後に加わる井村有希(ゆうき)といい、超一流のアマチュアの深い知識と熱意がこの研究の原動力となった。
並行して、全国の旧知のムシ屋や面識のない方々約50名へ材料採集の依頼状を送ったところ、次々とアルコール漬のオサムシが送られてきて、うれしい悲鳴をあげた。これらの方々の厚意に報いるため、新しい研究結果を逐次載せた「おさむしニュースレター」を、95年7月から99年3月20号まで、秘書・高橋弥生の手作りで発行した。
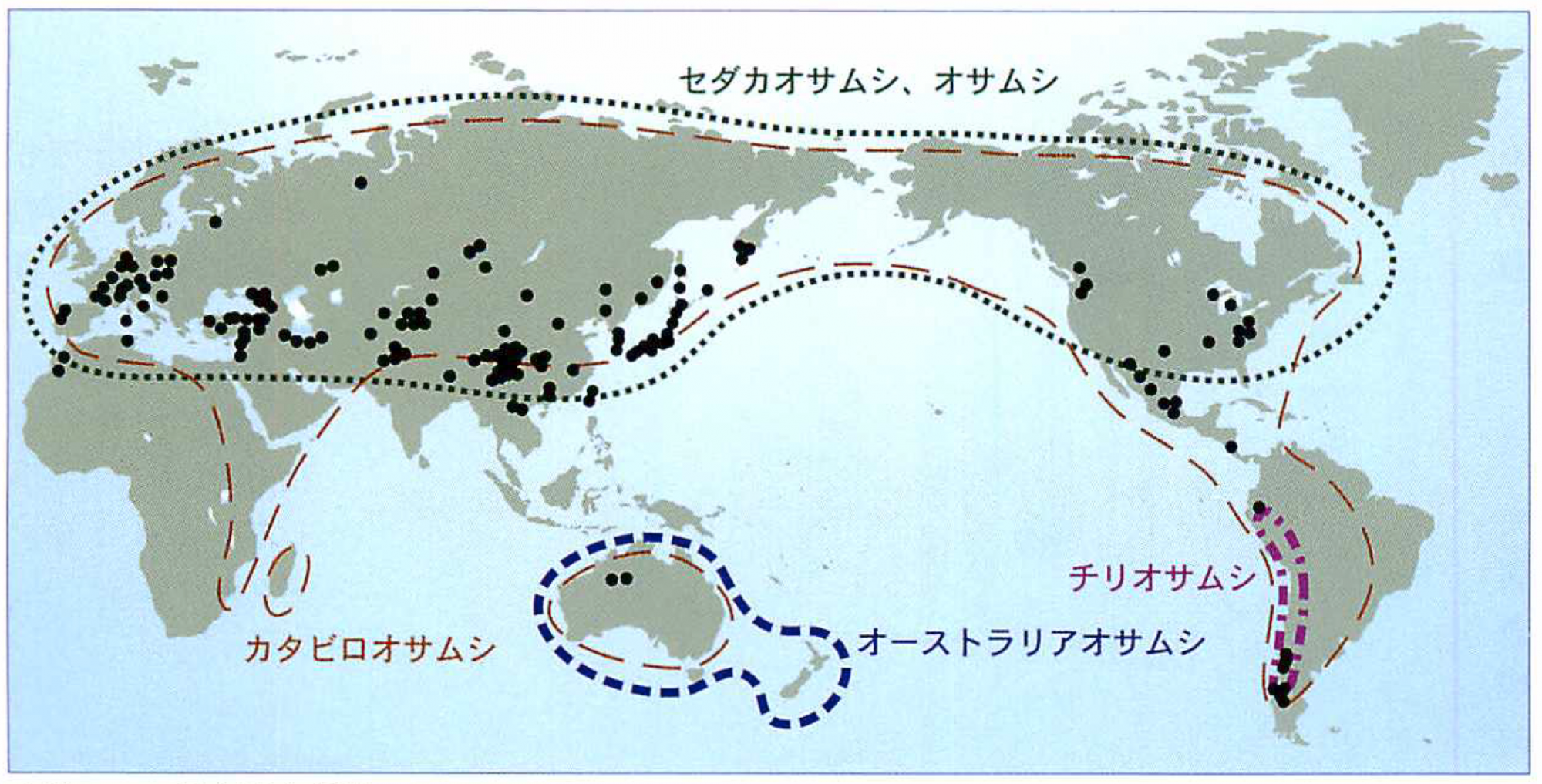
オサムシの分布域と採集地点
解析用のオサムシは世界中から集められ、約2000個体、350種を超えた。 黒丸は主な採集地点。
外へ広がる
95年3月、『科学朝日』の柏原精一が来訪、マイマイカブリとオオオサムシ属の解析結果が同誌に紹介された。その直後、姫路工業大学の松田高明から「マイマイカブリの分化から、日本列島形成が1500万年前と考えられるとのこと。古地磁気の測定でも同じ結果が出ているが」という電話を受けた。虫の研究と地球科学がつながったのだ。その後、共同研究者の乙藤洋一郎(神戸大学)と共に来訪され、古地磁気について教えてもらった。谷本周也(当時コミュニケーション部門スタッフ)が、地史とオサムシの分布に強い興味を持ち、情報を蒐集した。日本列島の地史とオサムシの分布に関する研究の基礎は、谷本と冨永が築いたといってよい。
オオオサムシ属の解析結果は、従来の分類学から見て余りにも"常識"外れで、ほとんど信用されなかったし、今でも信じない人は多い。形態の特徴によって分けられた4つのグループが、mtDNAで見ると地域的な5つの系統に入り交ったのだ。冨永はこの現象を「平行放散進化」、茂木和行(当時コミュニケーション部門ディレクター)は「不連続なタイプの変化、タイプスイッチングだ」と言う。半信半疑だったが、その後も同様の現象が多く見つかり、細かい点は別にしてタイプスイッチングによる平行放散進化のアイデアは誤りでないと考えている。
同年、日本鞘翅(しょうし)学会名古屋支部例会で結果を報告、総会の特別講演も行なった。話が終わるや池田清彦(山梨大学)が、オサムシの進化は、これまでの常識とは違うと思うがどう説明するのか、と聞くので、これこそ構造主義の出番ではないか、と切り返した。
池田には、後にオーストラリアオサムシの入手に尽力してもらった。また、この会で井村と会い、世界のオサムシ解析に向けて共同研究を組むことになった。もっともその頃の井村は、『世界のオサムシ大図鑑』完成直前で、せっかく形態からの分類がまとまりかけているのに、DNAという厄介なものがでてきたと、内心穏やかではなかったらしい。
世界のオサムシへ
95年の秋、岡田の口ききで、フランスから送られてきたオサムシ数種を蘇が分析し、日本のオサムシとともに系統樹に組むと、オサムシは、約4000万年前、40~50万年くらいの短期間に一斉に放散分化した(一斉放散)ことがわかった。
毎年チリで蝶採集の傍らオサムシも採っているという柏井伸夫は、見たこともないチリオサムシを次々届けてくれ、分類学的位置づけが不明確だったオーストラリアオサムシとチリオサムシの系統関係が決まった。チリオサムシについては、同所的平行進化という非常に興味ある事実も明るみに出たのである。海外のオサムシの解析は急速に進み出した。各国のオサムシ研究家と交流をもつ井村が、毎年欧州へ出かけ、膨大な数のオサムシを手に入れてくれたのだ。
また、中国科学院と共同で、井村、岡本、蘇が四川省で(97、98、99年)、97年からグループに加わった金衝坤(キムチュンゴン)と岡本、蘇が韓国で(98年)、採集調査をした。金は、短期間でヨロイオサムシ群に属する膨大な数のサンプルを精力的に分析し、オサムシの中で最も多様化の激しいこのグループの系統を明らかにした。
世界の約35カ国から集まった標本数は約2000、既知オサムシ族を構成する属の95%、総種数は350を越えた。オサムシの系統を語るに不足はない。
生命誌研究館でのDNA解析と平行して井村が詳細な形態学的研究を行い、2000年3月でほとんどの解析を終えた。形態学的研究では、いくつもの新種や新亜種が発見され、DNA解析ではオサムシの進化の様式や分布域形成に関する数々の興味ある結果が得られた。
新しい進化像
7年間にわたる研究の結果を要約すれば、第一に、オサムシの進化の過程で、大きな形態変化を伴うさまざまな規模の放散が起きたこと、別系統で同じような形態変化が起こる平行進化が見られること、長期間ほとんど形態が変わらない例もあること、が挙げられる。したがって進化は漸進的ではなく、「動」→「静」→「動」というように不連続に、二者択一的に起きると考えられる。この現象は、グールドら一部の古生物学者の主張とよく一致し、進化の姿を考察する上で興味深い。進化は「末広がり」という従来の見方とは逆に、総じて「先細り」のように見える。カンブリアの大爆発で動物の主要な門が一斉に出現したこと、オサムシの一斉放散とそれに続く中、小規模の放散などをみると、形態多様化の可塑性の範囲は、先細り的に狭まっていくようなのだ。同様のことは、グールドや神戸大学の河野和男も指摘している。遺伝子レベルでいうと、形態形成に関与する遺伝子群の発現が時間と共に制限されていくか、または遺伝子が消失していくかのいずれなのか、将来の検討が待たれる。
第二に、オサムシ相の形成が地史との関連で確立されたことが挙げられる。地球科学とのドッキングは、生物地理学に新しい光を投げかけた。
2000年2月、DNAによる系統解析の有効性をアピールし、新しい昆虫系統進化研究を推進する目的で、シンポジウム「DNA で辿る昆虫の系統と進化」を開催し、200名を越す参加を得た。オサムシを始めた当初は四面楚歌だったDNAも、ようやく二面楚歌あたりまでこぎつけたというのが実感である。
準備室時代、「生命誌研究館では日常性を持ちながら世界のトップクラスの研究がしたい」という中村に、「1研究室2~3人では無理」と言った。中村曰く「山椒は小粒でピリリと辛い」。私達のオサムシ研究がピリリと辛かったかどうかは第三者の判断にまかせよう。

世界のオサムシの系統解析を終え、形態の多様化から見えてきた進化のパターン。多様化の可塑性の「先細り」と同時に、進化の「静」と「動」を表している。オサムシ研究グループ(1999)。
(おおさわ・しょうぞう/JT生命誌研究館顧問)
※所属などはすべて季刊「生命誌」掲載当時の情報です。