RESEARCH
コケの細胞分裂にみる上陸の姿
生物の陸上進出といった大きな進化の背景には必ず、遺伝子の機能の変化がある。遺伝子、細胞、形づくりと、異なる階層の研究がつながって、陸上植物の進化の様相が見えてきた。
1.植物の陸上進出とコケ植物
私たちが暮らす陸の生態系は、植物によって支えられている。植物が上陸する以前の陸地は、岩石が剥き出しで乾燥し、昼夜の寒暖差が大きく、紫外線が降り注ぐという生物にとっては過酷な環境であった。もちろん従属栄養生物が生活するに足る栄養分も存在しない。荒れた大地にまず進出し、他の生きものが陸で暮らすための基盤になったという意味で、陸上植物の登場は生物史上の大きな出来事だったといえる。
最初に陸上に進出した植物がどのような姿をしていたかはわかっていない。姿がわかる大型化石で最も古いものは、ライニー植物群など枝分かれをした軸と胞子嚢を持つことが知られている。しかしより古い年代の地層から、現在のコケ植物に似た特徴を持つ胞子の化石が見つかっている。コケ植物のように小さく、厚いクチクラ層を持たない植物体は化石として残りにくいことから、初期の陸上植物は現在のコケ植物のような小さな姿をしていた可能性がある。
2.遺伝子の移動と陸上進出
陸上植物は、緑藻類のうち浅い淡水域に生育するストレプト藻類とよばれるグループから進化したと考えられている。陸上植物に最も近縁な藻類は、以前は、陸上植物を連想させる3次元の体をもつシャジクモ類だと考えられていたが、近年の大規模なゲノムデータを用いた分子系統解析から、「接合藻類」であることがわかってきた(図1)。接合藻類は、ミカヅキモのような単細胞からなるものや、アオミドロのように細胞が直線状に連なった単純な形を持つ藻類からなるグループである。その接合藻類は、GRAS遺伝子群(以降GRAS)という陸上植物と共通の遺伝子グループをもっていたのである。
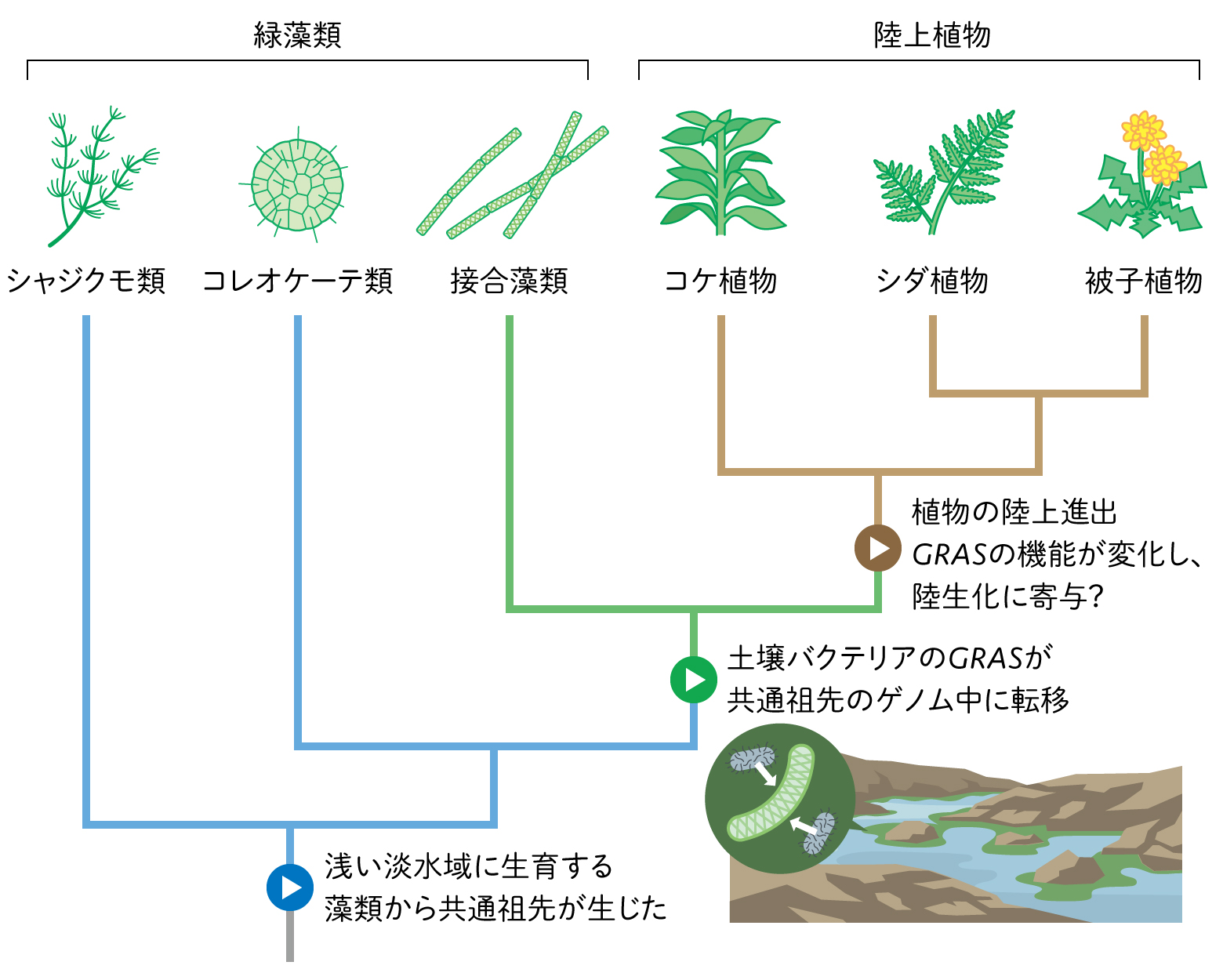
(図1)陸上植物の系統と遺伝子の移動
生きものの形が大きく変わるような進化の背景には、それに先んじた遺伝子の変化がある。細胞の中のゲノムは、生物が世代を重ねるうちに自ずと変化していく。これが進化の原動力となる。生物の進化の過程では、ゲノムが局所的に重複して遺伝子の数が増える現象や、ゲノム全体が重複して全ての遺伝子の数が倍になる現象が何回も起こり、増えた遺伝子からは新しい機能を持つものが現れる。ゲノムのDNA配列の解析技術や遺伝子の機能解析手法が進歩した現代では、そこに刻まれた歴史を調べることで、過去の出来事や祖先の姿により具体的に迫れるようになってきている。
最近の研究からGRASはバクテリアに由来する遺伝子であることがわかり、そこから、浅い淡水域で生育する藻類に土壌性のバクテリアが感染するなどして、そのバクテリアが持つGRASが接合藻類と陸上植物の共通祖先である藻類のゲノム中に転移したという道筋が提唱されている。さらに陸上植物の共通祖先の段階でGRASの遺伝子の数が増加し、陸上植物が多数のグループに分岐した後も大部分の遺伝子が引き継がれていることから、それらは陸上植物に共通する重要な性質をもたらした可能性がある。
バクテリアなど他の生物に由来する遺伝子には、乾燥ストレス耐性など植物の陸生化に貢献した重要な性質に関与するものが存在する。GRASは、植物の陸生化の際に何をもたらしたのだろう。陸上植物として最も種数が多く身近なグループは被子植物であり、遺伝子の機能についても最も研究が進んでいる。これによると、被子植物ではGRASの一つであるSHRという遺伝子が根ではたらいており、根の細胞分裂を制御して成長を促す働きや水を遮断する組織層を形成する働きを持つ。陸上生活において水を吸収し運搬する根の形づくりに、バクテリア由来のGRASが寄与している事実は興味深い。
3.植物の祖先の姿をコケに探る
コケ植物は陸上植物の進化の初期に、被子植物やシダ植物を含む維管束植物の共通祖先と分かれて独自に進化した系統であり、被子植物には受け継がれなかった祖先の性質を保持している可能性がある。上陸した祖先の姿をより明確に描くためには、被子植物の研究のみでは不可能で、コケ植物での知見との比較が不可欠である。
私たちは、コケ植物のヒメツリガネゴケを材料に用いている(図2)。このコケは、葉と茎からなる「茎葉体」という形をつくり、葉の真ん中には葉脈に似た水通導組織をもつ「中肋」と呼ばれる組織が作られる。GRASはシロイヌナズナでは根ではたらいていたが、根をもたないコケのGRASはどのような機能をもっているのだろうか。
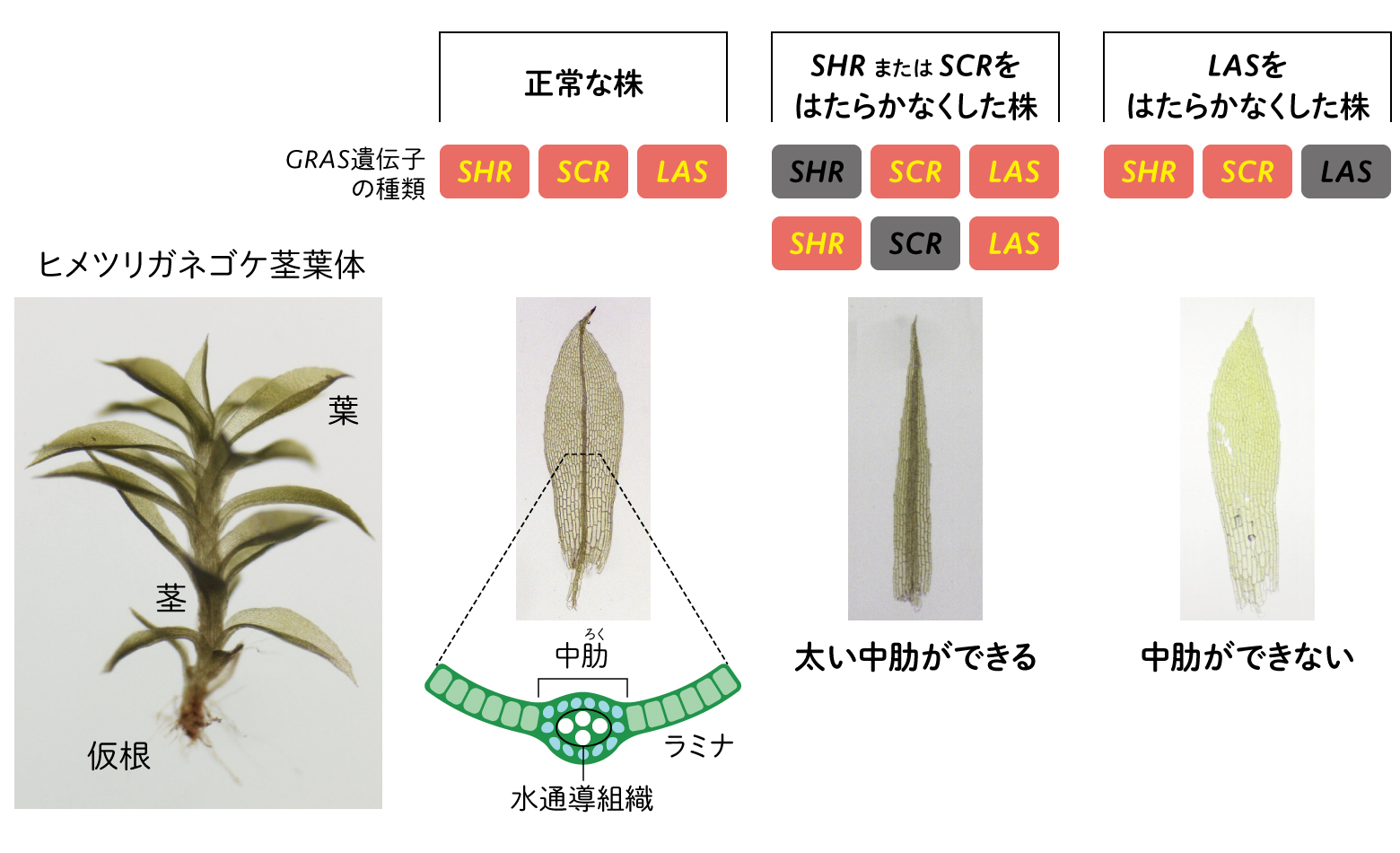
(図2)ヒメツリガネゴケにおけるGRAS遺伝子のはたらき
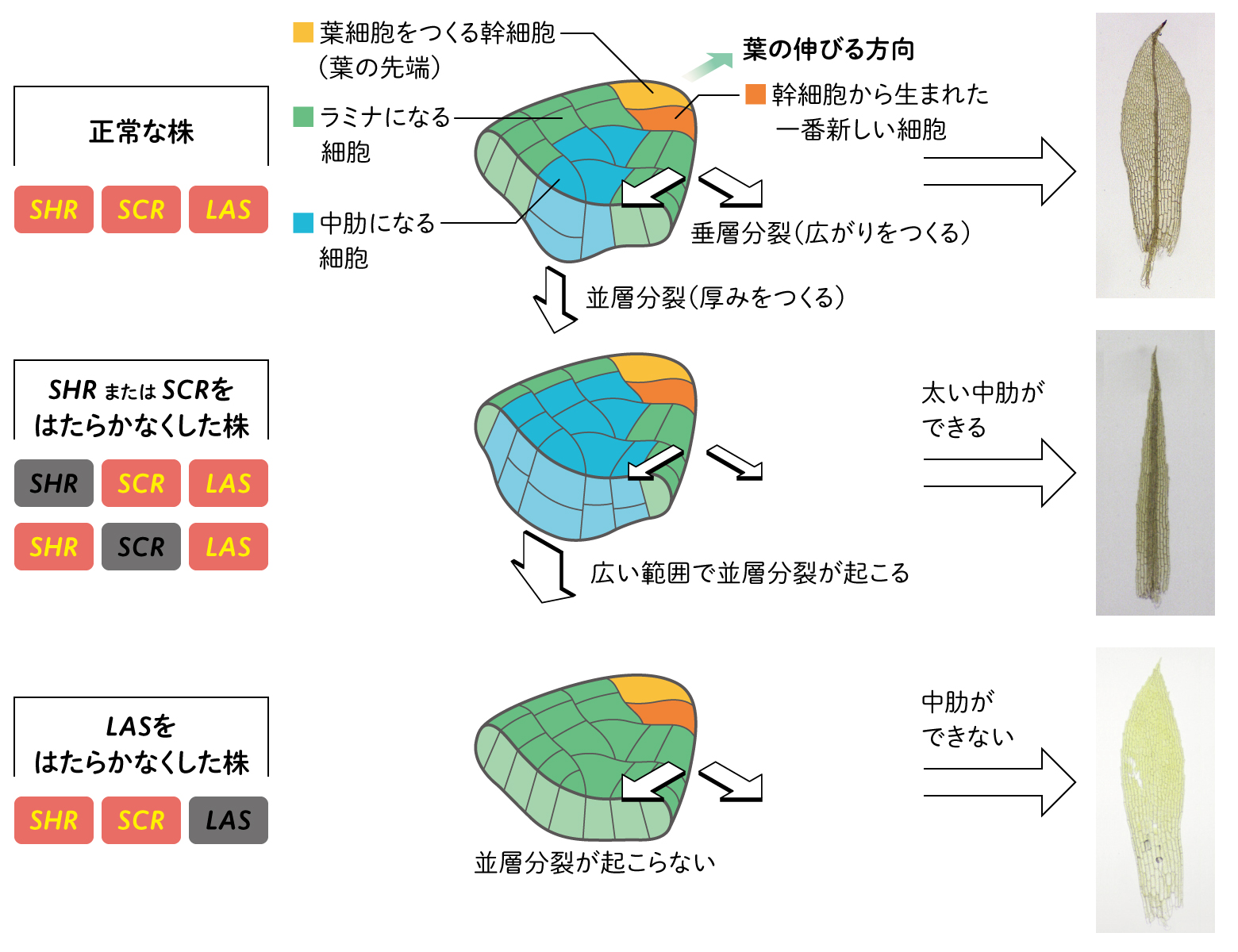
私たちは、ヒメツリガネゴケのGRASに属する遺伝子SHR、SCR、LASの3種類に着目した。これらの遺伝子の機能を順番に失わせてみると、極端に幅の広い中肋ができたり、逆に中肋が消失したりするなどの変化が見られた(図2)。やはりコケでもGRASは水通導組織の形成に関わっているようだ。
4.一つの幹細胞による葉の形づくり
植物の細胞は硬い細胞壁をもつため、生まれた細胞が移動することはできない。そのため細胞分裂の際に、どの方向に細胞を増やすかが後の形を大きく決定づける。被子植物は、茎の内部や枝の先、根の先に分裂組織をもっている。分裂組織には多数の幹細胞が存在しており、それらが、長さ方向や直径方向など、様々な方向に分裂することで立体的な組織をつくる。
一方ヒメツリガネゴケは、それぞれの器官はたった一つの幹細胞から作られる。一枚の葉をつくる際、茎の先端にあるたった一つの幹細胞から、中肋とラミナの両方を含む葉全体が作られる。役割の異なる組織が、1つの幹細胞からどのようにつくられるのかをまず観察しようと考えた。そこで、ヒメツリガネゴケの組織と細胞の一つひとつが、どのように分裂して形を作るかを、共焦点レーザー顕微鏡で詳細に観察した(図3)。
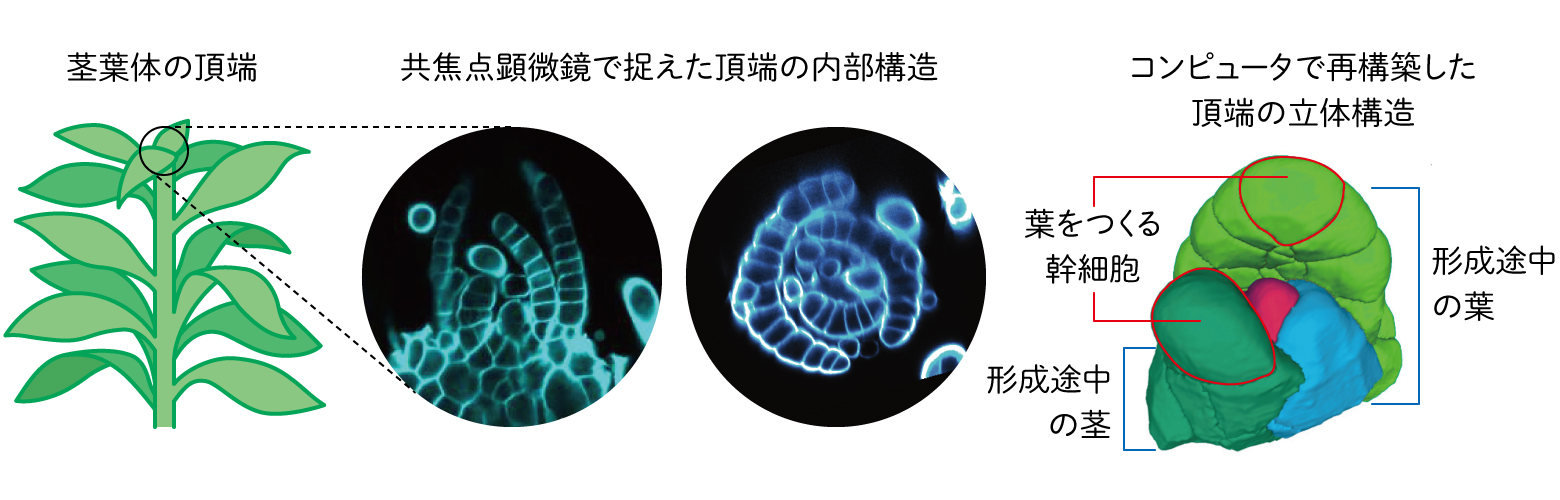
(図3)ヒメツリガネゴケの先端の幹細胞
ヒメツリガネゴケの葉の中肋は、水の通る細胞を、乾燥から守る表皮で包んだ細胞の束になっている。一方、葉の平面の部分であるラミナは全て一層の細胞からなる。ラミナをつくる細胞は、横に広がりをつくる方向に細胞分裂が進んでいく「垂層分裂」をするのに対し、中肋をつくる細胞は、縦に厚みをつくる方向に細胞分裂が進む「並層分裂」をする。
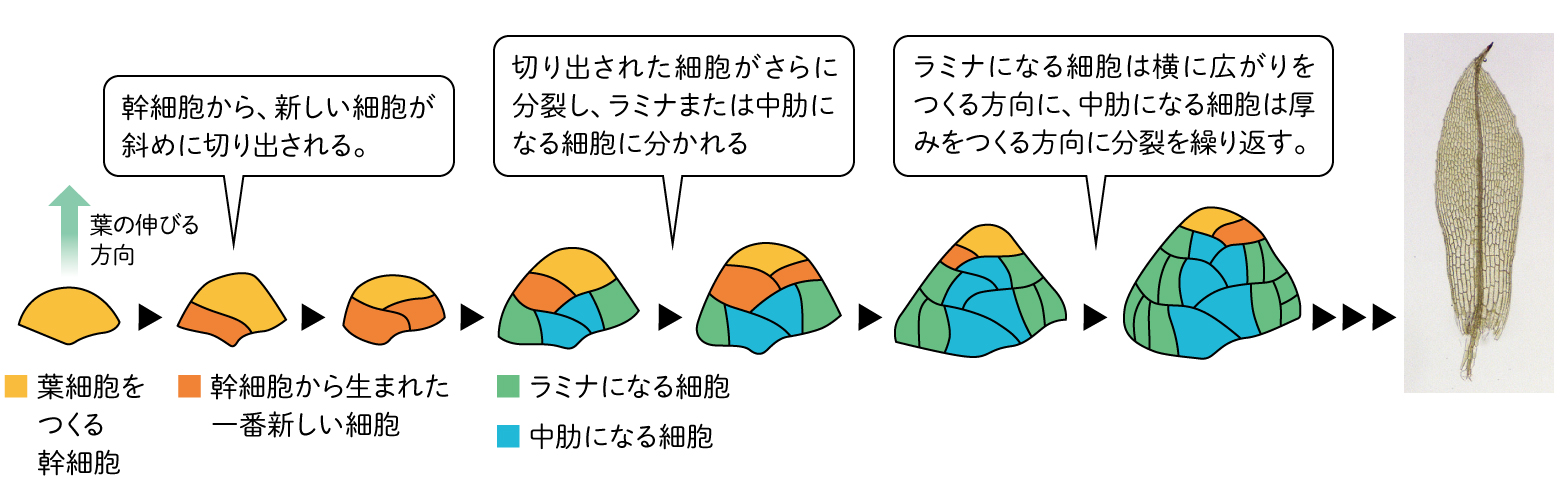
(図4)ヒメツリガネゴケの先端の幹細胞
5.細胞の形と分裂方向
一つひとつの細胞で、3種類のGRAS遺伝子と細胞分裂の関係を調べたところ、SCRあるいはSHRのはたらきをなくすと、正常な株より広い範囲で並層分裂が起こり、結果として中肋が太くなった。一方、LASのはたらきをなくすと並層分裂が抑制されて、中肋が消失することがわかった。
(図5)ヒメツリガネゴケの葉の形成とGRAS遺伝子の制御
3種類のGRAS遺伝子が、葉を作る細胞の分裂する方向を制御しているようだ。そこで3種類の遺伝子の関係を調べてみると、ラミナでは、SCRがSHRのはたらきを促進することで、厚み方向への分裂が抑制され、広がり方向への分裂が促進するのに対し、中肋では、LASがSHRを抑制することで、厚みを増す方向に細胞分裂が制御されていることが明らかになった。
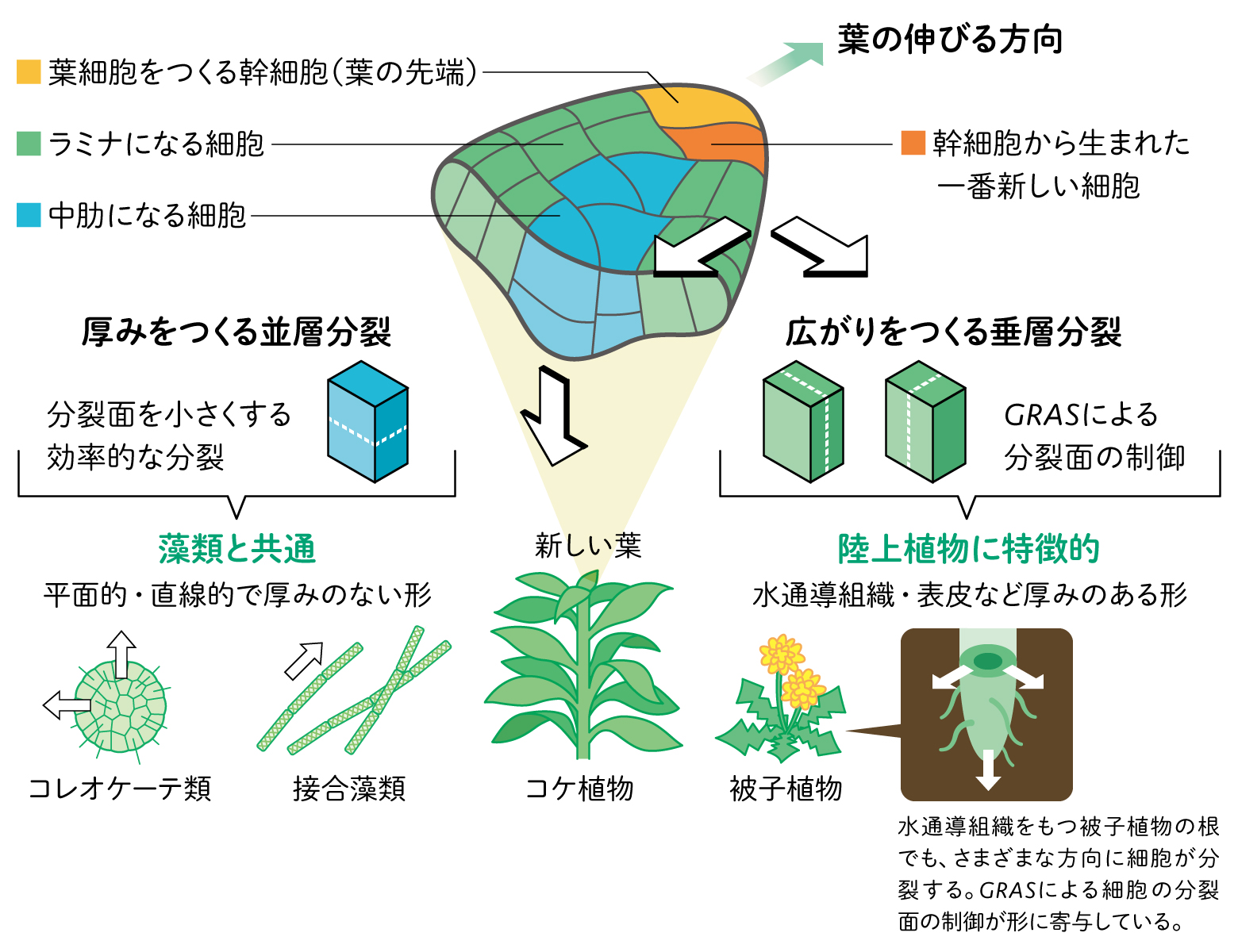
細胞の形をさらに詳細に解析してみると、ヒメツリガネゴケの並層分裂と垂層分裂では異なる点があることがわかった。それは細胞の分裂面の入り方である。
植物細胞が分裂する際、エネルギーや細胞壁などの資源を最も効率的に使って分裂するには、細胞の分裂面を小さくすることが有効である。緑藻類であるコレオケーテや接合藻類は、このように分裂する。同じ方向への分裂と伸長を繰り返すことになるため、結果として厚みのない平面的・直線的な形の多細胞体となる。
ヒメツリガネゴケの並層分裂では、分裂面を小さくする分裂が起こるのに対し、垂層分裂ではこれに従わず、葉の広がりをつくる方向に分裂面が入ることがわかった。複雑な器官をつくるには、分裂方向の制御が重要である。決まったタイミングで、決まった細胞を決まった方向に分裂させることが必須になる。GRASが特定の細胞の分裂面を特定の方向に制御するという、葉の形づくりのうえで重要な役割を担っていることが明らかになった。
藻類でこれまで行われてきた分裂と異なるタイプの細胞分裂がコケ類でみられ、さらにそれがGRASによって適切に制御されていることを発見した。陸上植物が複雑な器官形成をつくるしくみの一端を解明したと考えている。
(図6)細胞の分裂面と植物の形
6.陸への進化と形づくり
進化は起こったことの結果を後から解釈するしかないため、最初の陸上植物の本当の姿を知ることはできない。陸上植物は葉肉など水分の多い柔組織を乾燥から守るための、表皮をもつ。すなわち厚みをもった立体的な構造が陸上生活では必須となる。バクテリアから移動した遺伝子は多数発見されているが、大半は酵素などの遺伝子であり、陸上植物の立体的な形づくりの転写因子としてはたらいている遺伝子の発見は、GRASが初めてである。ここから、遺伝子ネットワークのどのような変化が、植物の陸上進出を可能にしたのか推測することができるだろう。
現在の植物科学の分野では、被子植物の研究が圧倒的に多い。これまで植物の細胞分裂を決める要因は、オーキシンなどのシグナル制御が重要だと考えられてきた。複雑なシグナル制御が成立する以前に、陸上植物がいかにして形をつくり始めたかについては謎だった。今回、細胞の形と分裂面の関係が、分裂方向の制御につながるという発見は、陸上植物の初期の進化から被子植物までを合わせて考える観点を生んだと考えている。

小藤 累美子(こふじ・るみこ)
金沢大学大学院自然科学研究科生命科学専攻博士課程修了。博士(理学)。基礎生物学研究所博士研究員を経て、2001年より金沢大学自然科学研究科助手。現在は金沢大学理工研究域生命理工学系・助教。

石川 雅樹(いしかわ・まさき)
名古屋大学大学院理学研究科生命理学専攻博士後期過程修了。博士(理学)。ロックフェラー大学博士研究員、ERATO博士研究員を経て、2012年より基礎生物学研究所・助教。














