RESEARCH
細胞記憶を支えるクロマチン
1.細胞記憶とは何か
私たち多細胞生物の体を形づくるすべての細胞は、ひとつの受精卵から受け継いだ同じゲノムDNAを持ち、はたらく遺伝子の違いによって多様な細胞へと分化する。いろいろな性質の細胞が全体として統制の取れた一つの個体になるには、細胞が分裂しても、もとの細胞の性質が正確に伝えられる必要がある。それが「細胞記憶」だ。細胞の性質は、はたらく遺伝子の組み合わせで決まり、それが安定して分裂後の細胞に伝わることが体の形づくりにはとても重要なのである。
たとえば、体節をつくるためにハエやマウスやヒトでもはたらいているHox遺伝子群は、胚発生初期に細胞ごとにはたらく遺伝子の組み合わせが決まり、それが細胞分裂後も長期にわたって維持されている。細胞の中でどの遺伝子がはたらくかが記憶されているのである。その結果、前の方から頭、胸、腹という順で形づくりが行われる。このような「細胞の記憶」、つまりはたらく遺伝子の組み合わせの維持は、どのように行われているのだろうか。
私たちは実験しやすく、遺伝学やDNAに関して多くの知見が蓄積しているショウジョウバエを用いて、細胞記憶の解明に挑んでいる。
2.遺伝子のはたらきかたを決定する因子
1930年に、マラーはショウジョウバエの赤眼の色を支配しているwhite(w)遺伝子座(註1)が、逆位(註2)によって、染色体のセントロメア(註3)近くに移動すると、赤白斑入りの眼が生じることを見出した。これは遺伝子が染色体のどこに位置するかによって、はたらいたりはたらかなかったりすることを示した古典的な研究で、この現象を斑入り位置効果(PEV:Position effect variegation)とよぶ(図1)。
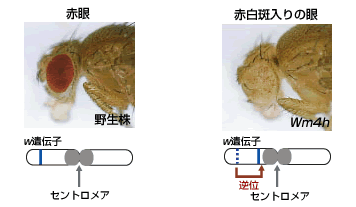
(図1) 斑入り位置効果のしくみ
Wm4h・・・white遺伝子がセントロメアの近くに転座したものの中で、特に遺伝子のはたらきが抑制されている変異株。
真核生物のゲノムDNAは染色体として核内に存在し、DNAとタンパク質が合わさったクロマチン繊維が何重にも巻いた複雑な構造をしている(図2)。染色体全体にはクロマチン繊維が凝集して遺伝子があってもはたらかない領域(ヘテロクロマチン)と、ほどけていて遺伝子が活発にはたらく領域(ユークロマチン)とがある。染色体末端のテロメアやセントロメア付近は、ヘテロクロマチン領域になっている。斑入り位置効果はw遺伝子座がセントロメア近くのヘテロクロマチン領域に移動したことで、w遺伝子がはたらく細胞とはたらかない細胞ができ、それが記憶となって分裂後の細胞に伝達されるために起きた現象である(図3)。このような位置効果と共に重要な役割を果たすのが遺伝子を活性化したり、抑制したりする因子である。
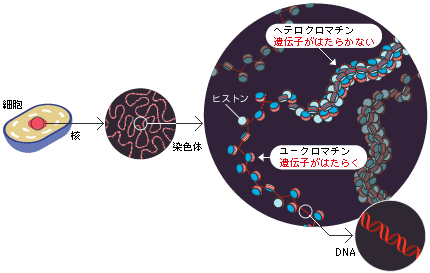
(図2) クロマチン繊維の構造
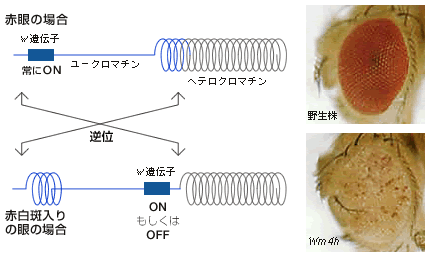
(図3)斑入り位置効果とクロマチンの関係
冒頭で述べたHox遺伝子群がどのような組み合わせではたらくかは、遺伝子のはたらきを抑制するポリコーム遺伝子群と、活性化するトリソラックス遺伝子群とが拮抗することで決まる。斑入り位置効果でも、遺伝子のはたらきを抑制する因子と活性化する因子とが、多数報告されている。図4で示す通り、Hox遺伝子群と斑入り位置効果では、それぞれに遺伝子のはたらきを制御する因子があり、その多くは異なっているがGAGA因子をつくるTrithorax-like(Trl)遺伝子だけは両方に関わっている。私たちはこれが共通するしくみを探る鍵になりそうだと考え、GAGA因子を手がかりに遺伝子のはたらきがどのように調節されるかを調べた。
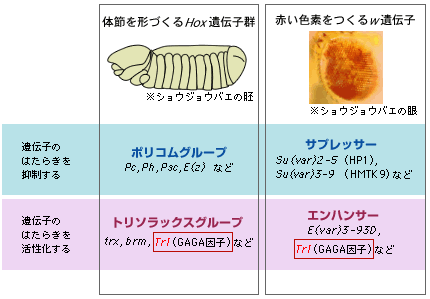
(図4) 遺伝子のはたらきを抑制する因子と活性化する因子
(註1) 遺伝子座
染色体上での遺伝子の位置、また遺伝子の領域が占める染色体上での区画を意味する。
(註2)逆位
染色体中にある一部のDNA配列が、本来の向きとは逆転した状態で存在していること。
(註3)セントロメア
染色体の長腕と短腕が交差する部位で、染色体のほぼ中央に位置している。セントロメアのクロマチンは高度に濃縮されている。
3.GAGA因子と結合するFACTの発見
GAGA因子はGAGAGという塩基配列を識別して、DNAに結合するタンパク質である。ショウジョウバエでは眼の色に関わるw遺伝子も含め、さまざまな遺伝子でそのはたらきを制御する領域にGAGA因子の結合配列が集合している場合が多い。凝集したクロマチンをほどく反応をクロマチン構造変動とよぶが、GAGA因子は結合配列の近くでクロマチン構造を変動させることがわかっている。しかしこの因子だけでは、クロマチン構造を変動させるには不足であることもわかってきた。ここで必要なもう一つの因子はGAGA因子と相互作用するタンパク質に違いないと考え、その探索を進めた。
考え方は決まったが、そのようなタンパク質を探すには一体どうしたらよいのか、そこから始めなければならない。そこで培養細胞で用いられている方法を個体に適用してみようと考えた(図5)。
まず、Flagという目印のついたGAGA因子(以下Flag-GAGA因子)の遺伝子を入れたトランスジェニックショウジョウバエの胚を作製した。この胚の細胞内ではFlag-GAGA因子がつくられ、そこには相互作用するタンパク質も結合しているはずである。この胚の核抽出液からFlag抗体ビーズを使ってタンパク質を集めた。Flag-GAGA因子とそれに結合している未知のタンパク質がここにあるはずである。個体発生の過程でダイナミックに活動している細胞でこの方法を試した例はなかったが、幸いうまくいき、GAGA因子と相互作用するタンパク質としてSSRP1 (Structure-Specific Recognition Protein 1)とSPT16 (Suppressor of Ty 16)を同定した。
SSRP1とSPT16は、酵母からヒトまで真核生物に広く保存されており、FACT (facilitates chromatin transcription)とよばれるヘテロダイマーを形成するタンパク質として知られている。FACTは、DNAからRNAが合成される際にヌクレオソーム(註4)を変形させてRNAポリメラーゼIIの通過を助けるとともにクロマチン構造変動を促進するという2つの働きがある。私たちが見つけたタンパク質は、まさに求めていたものであると考えてよい。次に私たちは、このタンパク質がクロマチン構造変動にどう関わるのかに注目して実験を進めた。
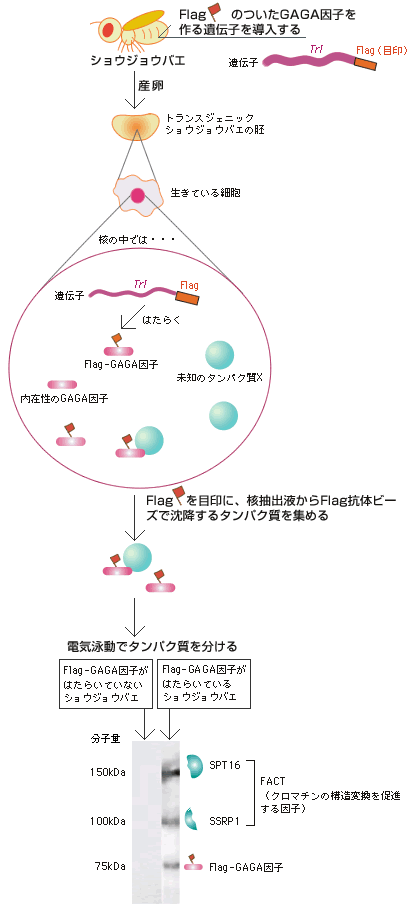
(図5) Flag-GAGA因子に結合するFACTの検出方法
(註4)ヌクレオソーム
クロマチンの基本構造。ヒストンタンパク質にDNAが1.75回転巻き付いている(図7参照)
4.GAGA-FACT複合体がクロマチンの構造変動をおこす
GAGA-FACT複合体が、斑入り位置効果に関わる因子であるかどうかを確かめるために、まずいくつかの遺伝学的解析を行った。染色体の逆位によってw遺伝子がセントロメアの近くに転座した変異株(wm4)では、その表現型として眼の一部に白い斑入りを生じることはすでに述べた(図6)。
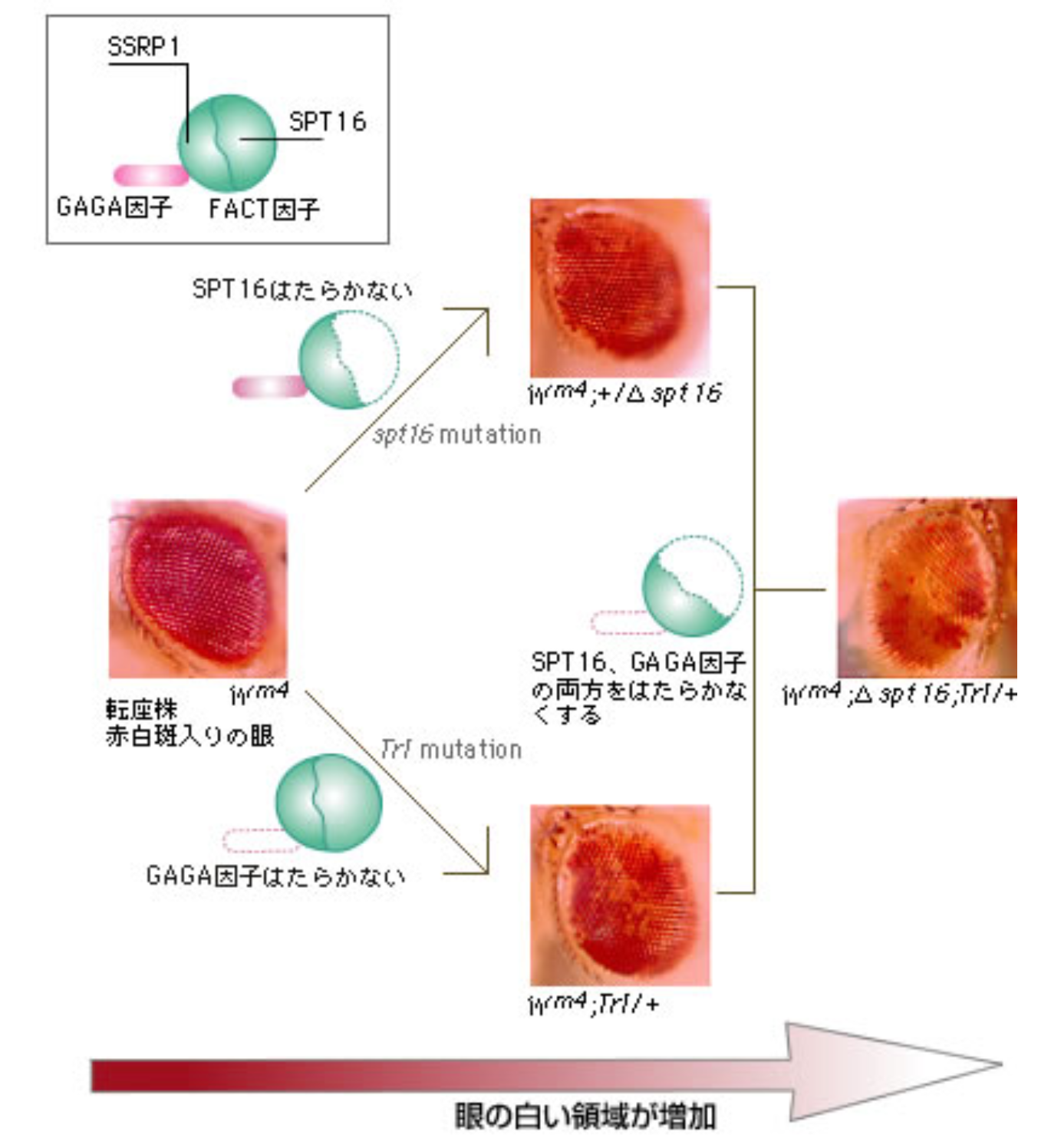
(図6) GAGA-FACT複合体と斑入り位置効果の関係
この変異株で、GAGA因子をつくるTrl遺伝子もしくはFACTの一部であるSPT16をつくる spt16遺伝子をはたらかないようにしたところ、眼の中の白い領域が増えた。次にTrl遺伝子とspt16遺伝子の両方をはたらかないようにしたら、さらに白い領域が増えた。これらの結果からGAGA-FACT複合体が斑入り位置効果に影響を与えているに違いないことが見えてきた。そこで、この変異株のw遺伝子座の近くを調べたところ、w遺伝子座とセントロメアヘテロクロマチン領域との間に位置するGAGAG配列(以下GAGA因子結合配列)上にGAGA-FACT複合体が検出された。GAGA因子結合配列は、GAGA-FACT複合体が結合することによって、DNAを切断するヌクレアーゼへの感受性が高まることもわかった。ヌクレアーゼは、凝集したクロマチン上でははたらかないので、この領域でGAGA-FACT複合体によるクロマチン構造変動が起きていると推測できる。複合体がクロマチン構造変動をおこすメカニズム、次の問いはこれである。
5.ヘテロクロマチンに対する防波堤を築く
DNAはヒストンに巻き付いた状態で核の中に収納されている。ヒストンとは、H2A, H2B, H3, H4のタンパク質がぎっしり詰まった8量体タンパク質である(図7)。
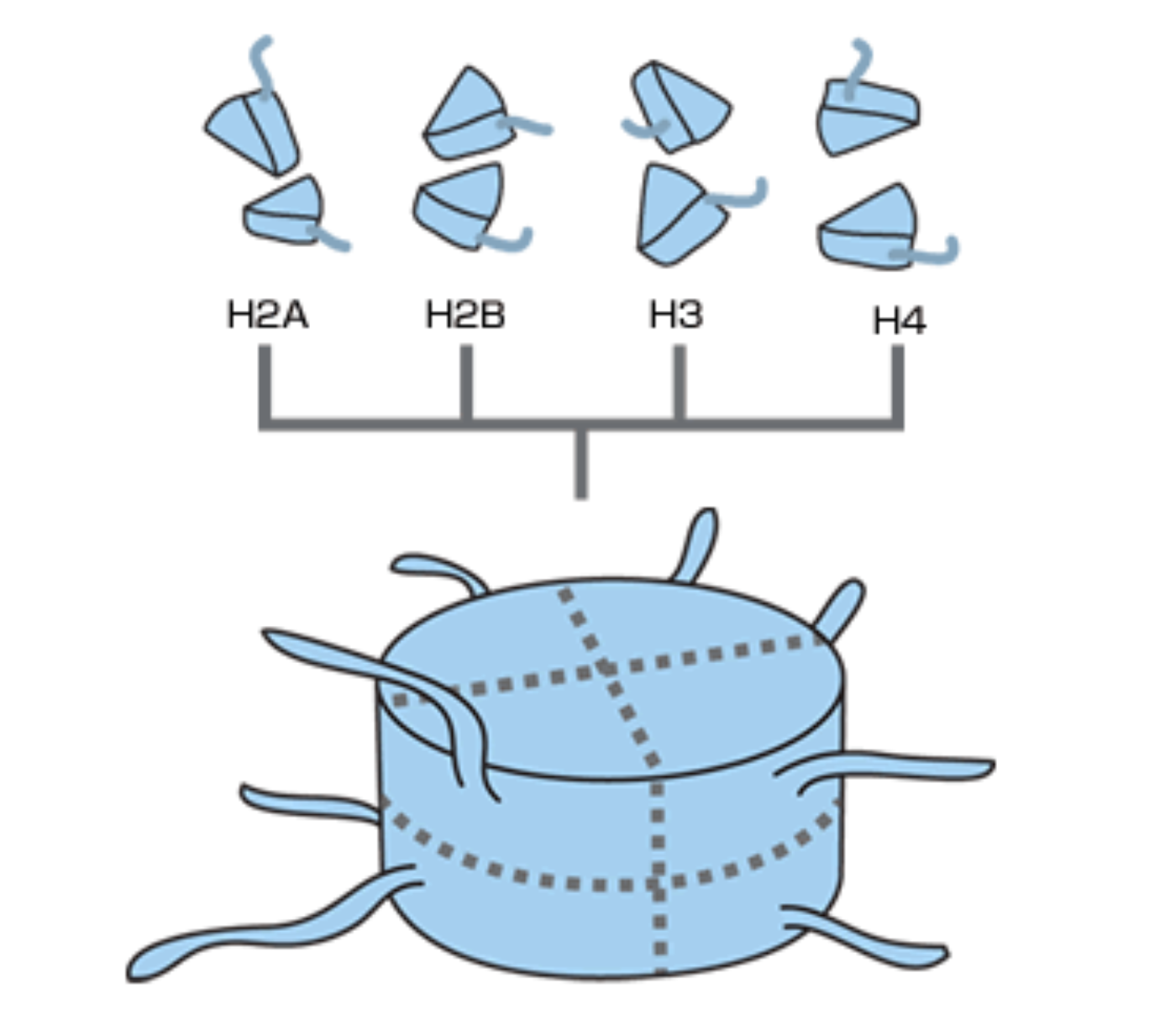
(図7) ヒストンの構造としっぽ
最近の研究からヒストンは、単なる “糸巻きの芯”ではなく、しっぽのように突き出した部分にあるアミノ酸が、メチル化(註5)などの修飾を受けることによってさまざまなタンパク質と相互作用し、遺伝子のはたらきにも積極的に関わっていることがわかってきた。
例えばヒストンH3のしっぽにある9番目のリジンのメチル化(以下K9メチルH3とよぶ)がクロマチンの凝集に関わっており、遺伝子がはたらいていない目印になっている。この抑制性の目印を指標として、GAGA-FACT複合体のクロマチン凝集への影響を調べた(図8)。
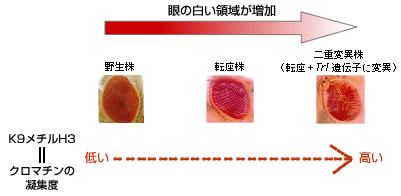
(図8) GAGA-FACT複合体のヒストンメチル化への影響
その結果、染色体の逆位が起きていない野生株のw遺伝子座では、クロマチンが凝集している状態を示すK9メチルH3がほとんど検出されないのに対し、w遺伝子転座変異株では少し検出されたのである。さらにw遺伝子の転座に加えてGAGA因子をつくるTrl遺伝子がはたらかない二重変異株では、メチル化は通常のセントロメアのヘテロクロマチン領域と同程度に高いレベルで検出された。この変異株ではTrl遺伝子がはたらかなくなったことでGAGA因子がつくられず、GAGA-FACT複合体が正常に機能しなくなったためにw遺伝子を含む領域が凝集し、ヘテロクロマチン化したのだろう。
以上の結果から、GAGA因子結合配列に結合したGAGA-FACT複合体は、セントロメアからw遺伝子座へのヘテロクロマチンの侵攻をせき止める防波堤の役割をしていることが推測できる(図9)。つまりGAGA-FACT複合体が結合している場所を境にしたヘテロクロマチンの逆側の領域は、クロマチンをほどけた状態に保っているので、遺伝子がはたらけるのである。この複合体が結合していない場合はヘテロクロマチンが侵攻して遺伝子がはたらかなくなってしまうのだろう。私たちは、GAGA-FACT複合体が斑入り位置効果と同様のしくみでHox遺伝子群のはたらきを維持していることも明らかにしている。
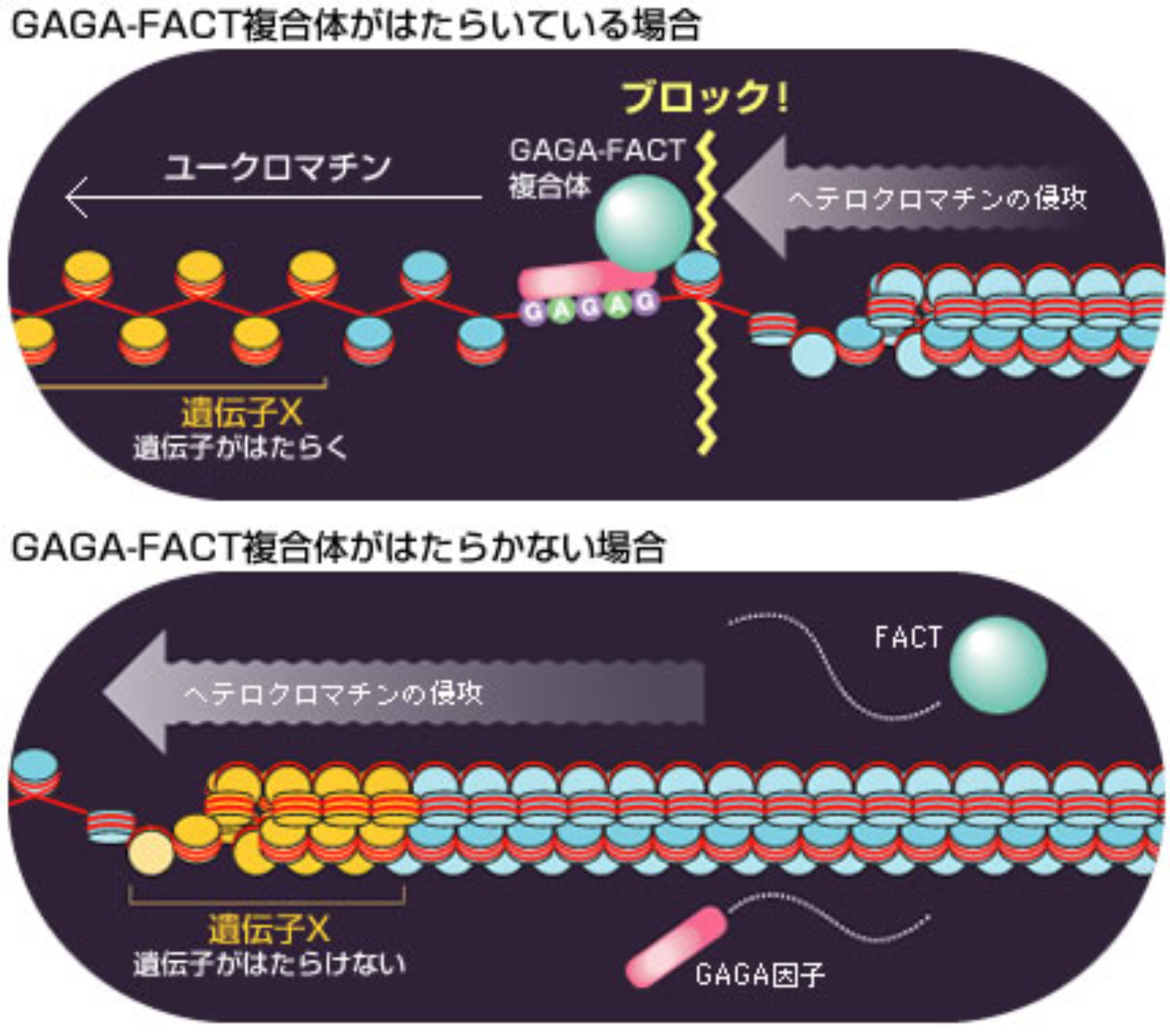
(図9) 防波堤としてはたらくGAGA-FACT複合体
さらにGAGA因子結合配列付近を詳細に調べたところ、そこではヒストンH3が未修飾ヒストンH3.3に置換されていることが分かった。GAGA-FACT複合体が抑制性の目印のついたヒストンH3を異性体である未修飾ヒストンH3.3に交換することによってヘテロクロマチンの侵攻を食い止めていることが見えてきたのである(図10)。H3.3は核内でも転写がさかんに行われている領域に局在しているヒストンなので、これが凝集するクロマチンに対する防波堤の中核を担っているのではないかと考えている。
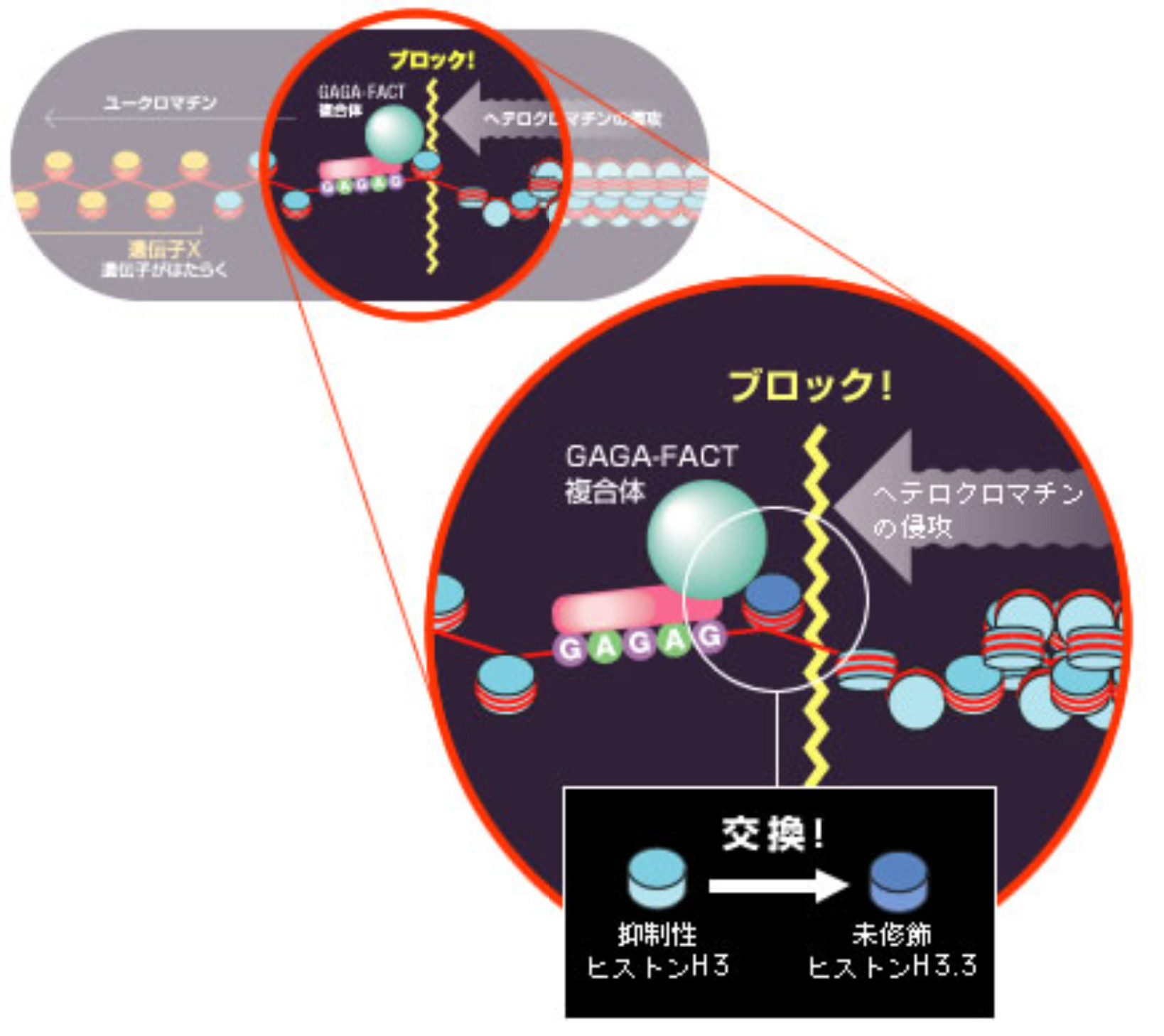
(図10)抑制性ヒストンH3から未修飾のヒストン異性体H3.3への交換
(註5) メチル化
タンパク質のアミノ酸側鎖にメチル基が結合する反応。ヒストンのしっぽの部分のアミノ酸が修飾されることによって遺伝子のはたらきを制御している。
6.「記憶」されるクロマチンの状態
これまでの結果を総合して、私たちは現在「細胞記憶」について以下のモデルを想定している。ヘテロクロマチンではヒストンH3の9番目や27番目のリジンがメチル化されており、これを目印にしてヘテロクロマチン化の役割をするタンパク質HP-1や不活性クロマチン状態を維持するポリコームタンパク質群がDNAに結合する。ヘテロクロマチンは本来周囲に広がる性質を持っているのだが、GAGA因子結合配列にGAGA-FACT複合体が結合するとクロマチン構造変動が引き起こされ、複合体が結合した場所にある抑制性ヒストンH3が、未修飾ヒストンH3.3に置き換えられる(図10)。こうしてヘテロクロマチンの侵攻を防ぐのでその先の遺伝子ははたらけるのである。またGAGA-FACT複合体はゲノム中にあるGAGA因子結合配列全てに結合するわけではなく、ある特定のGAGA因子結合配列に結合することで遺伝子のはたらき方を制御し、細胞の性質を決めている。さらに細胞分裂後もこの複合体の結合は保たれるので遺伝子ははたらき続け、それぞれの特徴をもつ細胞が生まれ、組織、器官をつくっていくのである。
GAGA-FACT複合体は、クロマチンの状態を巧みに変化させることで遺伝子のはたらきを制御し、それを娘細胞に継承していくことによって個体の形づくりを支えているのである(図11)。現在では分子についてもさまざまな目印をつけてその動きを捉える、別の表現をするなら“観る”ことができるようになった。これが生命現象の具体的な理解を助けている。クロマチンのダイナミックな状態変化の解析を基本にして、「遺伝子のはたらき方」を細胞が「記憶」する様子を調べ、生きものの形づくりを知るための研究を続けていきたい。
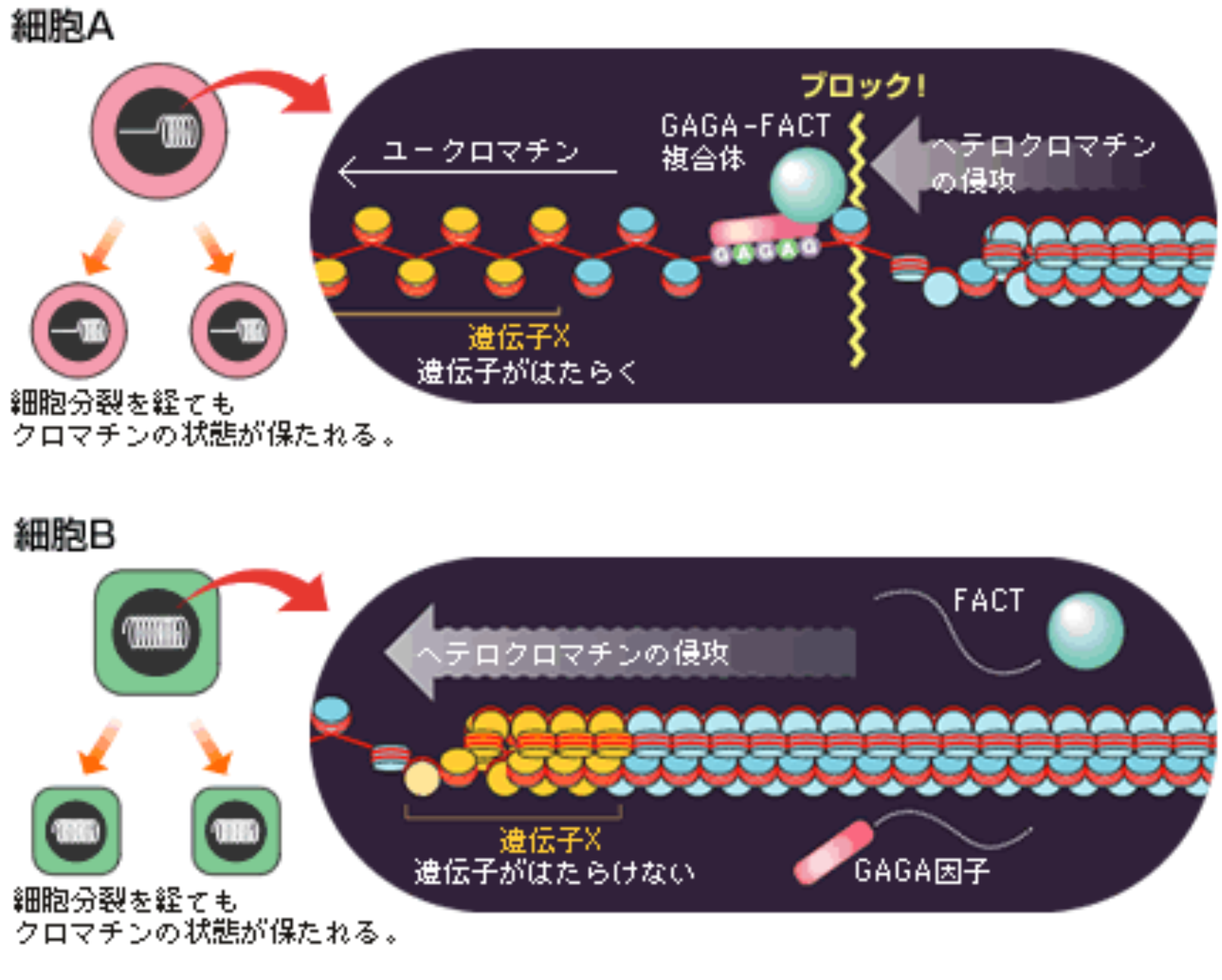
(図11) 記憶されるクロマチンの状態
広瀬 進(ひろせ・すすむ)
1943年生まれ。東京大学大学院理学研究科修了、理学博士。名古屋大学理学部助手、基礎生物学研究所助教授を経て現在国立遺伝学研究所教授で副所長。総合研究大学院大学生命科学研究科教授併任。














