News & Views
平行放散進化 -多様化の新しい原理
生物はどのようにして進化・多様化してきたのだろうか。 JT生命誌研究館で進めているDNAによるオサムシの系統解析は、「平行放散進化」と名づけられる新しい多様化のメカニズムの存在を示唆し、 種の概念の再検討を迫っている。
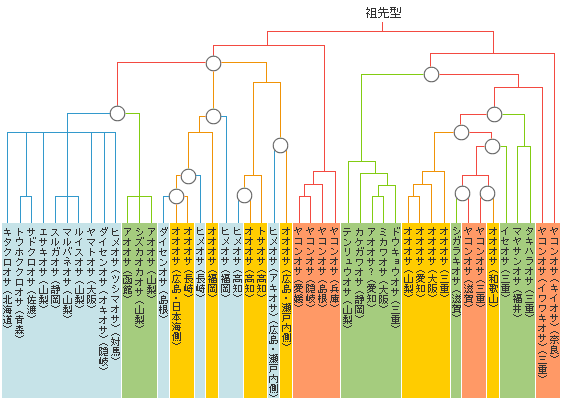
JT生命誌研究館では、 日本産のオサムシ全種の系統図づくりを蘇智慧(スーズィフィ)研究員のもとで1994年4月から進めてきた。オサムシは少数の例外を除いて翅が退化して飛べないので、地理的な分布が種の多様性を反映していることが多く、進化の系統解析に向いている。元東京都立大の石川良輔が、形態分類から一歩突っ込んで交尾片による分類を試み、その形の変遷から一種の定向進化を導き出した。 その検証を遺伝子レベルで進めようというのが当初の狙いだったが、ミトコンドリアのND5遺伝子は思わぬ事実をさし示した。
ミトコンドリア遺伝子によるオサムシの系統樹は、おおむね形態による分類関係を反映しているが 1)、オオオサムシ、ヤコンオサムシ、ヒメオサムシの3種は、さまざまな系統へと分散し形態による分類関係をこわしている。オオオサムシは、黒青色の大型のオサムシで、中部から近畿、中国、四国、九州に分布している。本来なら系統樹上で一つのまとまりとして出るはずなのに、もっと小型で形態の異なるほかのオサムシのおよそ4つのグループへと放散してしまう。1968年に、鳥取県の辰巳峠で見つかったオサムシの翅の化石(→右図)(900万~1000万年前)を基準として系統樹にあてはめると、和歌山と四国のオオオサムシは少なくとも数百万年も前に分岐した遠い系統に入る。ヤコンオサムシについても、2つの遠く離れた系統に分かれてしまう。ヒメオサムシとその近縁(亜)種でも事情は同じである。

【オサムシの翅化石】
900万~1000万年前。鳥取県辰巳峠産。 (大阪市立自然史博物館蔵)
【平行放散進化を示すオサムシ】

(左)【オオオサムシ】写真=松香宏隆 (中央)【ヤコンオサムシ】写真=冨永修 (右)【ヒメオサムシ】写真=松香宏隆
遺伝子によるオサムシの系統樹づくりを指導してきた生命誌研究館顧問の大澤省三は、この現象を「平行放散進化」(Parallel Radiation in Evolution)と名づけた1)。通常「平行進化」とは、異なる進化系列の生物が、たがいによく似た形態と機能をもつように進化していく現象をいう。新大陸の有袋類と旧大陸の真獣類とが、生活環境・生活様式の類似性からよく似た形へと別々に進化したのはその好例である。この種の収斂 現象は適応が形を生む、と説明される。しかし、今回の事例は、むしろ「形態の特長を支配する複数の遺伝子を発現させる上位の遺伝子が、それぞれの地域で独立に同様な変異を起こすことによって、形態的に区別 のつかないオサムシが平行進化した結果である、と考えたほうが自然ではないか」と大澤は言う。
こうした形態の独立的な変異を、大澤は「タイプ・スイッチング」と名づけた。よく似た平行放散進化の現象が南米のドクチョウ(→右図)で知られている。形態による系統では同じ種に入るものが、ミトコンドリア遺伝子による系統解析の結果 、いくつもの離れた系統に放散してしまう(ブラウアー、1994[下図) 3)。この一帯では、氷河時代の激しい気候変動によって、地理的な飛び地がいくつもできており、その飛び地のそれぞれで同一の紋様が独立に進化したように見える。昔からよく知られていたショウジョウバエの射精管の分化もよく似た現象である。これもまた、まったく違った進化系列に同じ形の射精管が現れる(スロックモルトン、1965)4)。

【南米のドクチョウ】
(写真=山口進)
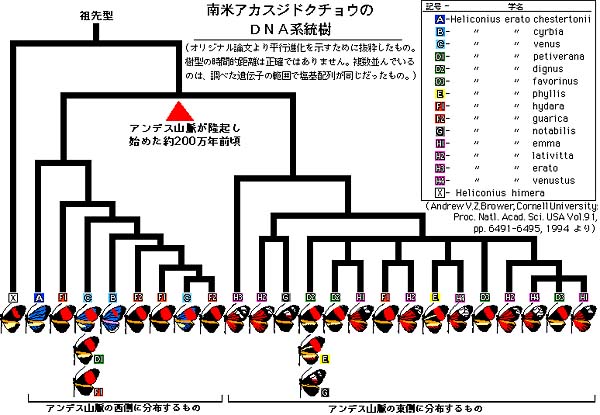
ブラウアー、1994
カンブリア紀の大爆発(5億年前)や昆虫の爆発的な多様化(3~4億年前)、恐竜時代の終焉後に起きた哺乳類の多様化(6000万年前)など、進化史上では種が一気に多様化する現象がいくつも知られている。スミソニアンのアーウィンは、熱帯雨林の研究から、1本の木には昆虫を主体とする600種の節足動物がいることを見出した。5万種の樹木があることを考えると、熱帯雨林だけで3000万種もの昆虫類がいることになるという(1982)5)。爆発的な多様化や、限られた地域における種の密集という事実は、「ダーウィン流の自然淘汰の結果とするよりも、タイプ・スイッチングなどによって、さまざまな形が一気に生み出され放散した、と考えたほうが合理的なのではないか」と、大澤は考える。
タイプスイッチングと平行放散進化
「放散」進化のアイデアを出したのは、オサムシの資料提供を熱心に進めたアマチュア昆虫研究家の冨永修である。冨永は仲間と近畿オサムシ研究グループを作り、近畿地方のオサムシの分布図を作ろうと20年以上前から進めていた。この一帯には、マヤサンオサムシやイワワキオサムシ、ヤコンオサムシなど6種類が分布しているが、オオオサムシの分布を調べるうちにいくつかの奇妙な空白域の存在に気づいた。「オオオサムシが、ある系統から均一に進化して拡散していったとすると、古地理の変化を考慮に入れたとしても空白域が説明できない。遺伝子の解析結果 を見てなるほどと思った。それぞれの地域で独立して同じ形態が分散して進化したとすれば よく説明がつく。そこで、これはいろいろなタイプが独立してバーッと出てくることになるのだから、放散進化ではないか、と言ったのです」と、冨永は語る。
自然淘汰説に対して故木村資生(きむらもとお)は、分子進化の中立説をたて、種の多様化の原動力として中立的な遺伝子の偶然の浮動をおいた 6)。タイプ・スイッチング仮説は、表現型に反映した中立的な遺伝子の変化が、自然淘汰的な圧力を受けずに集団へと広がっていく可能性を示唆しているのではないか。

アワフキムシ。
自然淘汰よりも中立進化で多様化した可能性が見られる。
宮田隆(京都大学、分子進化学)は、多種類の遺伝子の進化を調べ、力ンブリア紀には分子レベルでの中立的な変異が非常に多いことを見いだした。宮田によると、この時期に自然淘汰圧がゆるみ、普通なら有害で淘汰されてしまうような変異が中立となって生物の形態の多様化が促進されたのではないか、と言う(1994)7)。オサムシの平行放散進化の例は、中立的変異の蓄積→形態の多様化、といったシナリオが、必ずしも自然淘汰の有無にかかわらず、進化のさまざまな局面でかなり常態的に起きていることを示しているのではないか、と大澤は言う。
オサムシのなかでも、オオオサムシ亜属の仲間は、交尾器に付随したキチン質の骨片(交尾片)の多様なことで知られている。そして雌にはそれに対応した腟盲嚢の存在が知られており(石川、1973)8)、鍵と鍵穴の関係で生殖的隔離が行なわれる明確な例とされる。実際、交尾器が違いすぎるオサムシ同士が交尾すると、腟に出血を起こして雌が死んでしまったり、雄の交尾片が折れてしまうといった結果が多数観察されている(曾田・久保田、1995)9)。石川良輔はこの交尾片を分類の指標に使い、三角型、桃の実型、フック型などオオオサムシ亜属を5つの種群に分けた。そして、祖先型からこの交尾片がそれぞれが小さいほう(単純)から大きいほう(複雑)へと定向的に進化したと考えたのだった。しかし、今回のミトコンドリア遺伝子による系統解析は、この考え方も支持しない。異なったタイプの交尾片が分岐上では同じグループに入ったり、逆に同じタイプの交尾片が系統樹上では遠くに離れたりしてしまう。むしろ、交尾片の分化も、タイプ・スイッチング的な中立的多様化であることをミトコンドリアDNAの解析は示唆している。
交尾器の鍵と鍵穴の関係は、必ずしも機能していない、という実験がジャノメチョウである(グールソン、l993)。グールソン 10)によれば、分類学者が交尾器の違いにあげていた形質は現実には交尾で使われていない場所だった。オサムシでも、「鍵」は必ずしも完全ではない、との研究が出てきている(曾田・久保田、1995)9)。シロチョウの仲間では、交尾器の形の違う種間で人工的に交雑させると、F2までは雑種が育つことが調べられている(ポーター&シャピロ、1990)11)。 グールソンは、交尾器の変異は自然選択によるだけでなく、中立的な分化の可能性もあることを指摘した(1993)10)。北海道大学の小松正(昆虫体系学)は、交尾器の進化は、中立説・自然選択説のどららを支持するか、アワフキムシを使って検証しようと試みた。
小松は交尾器で8形質、全身形態で10形質を選び出し、札幌市内の7つの地域で採集したポプラなどにつくシロオビアワフキについて、個体群内および個体群間の分散と相関を調べた 12)。また、シロオビアワフキとその近縁種の5種について、同じ形質の分散と相関を調べた。その結果、交尾器の形態は個体サイズの大小にかかわらず種内変異が大きくなり、種内変異は小さいとする自然選択説による予測と逆の結果が出た。これは種間の変異にもあてはまり、むしろ偶然の浮動によって形が多様化するという中立説を支持している、と小松は言う。ただし、自然選択をゼロと仮定したときの理論的種間分散よりも得られた結果は小さすぎるので、不利な変異を除く負の自然選択などによってこの多様化は制限されていると見るべきだ、とも小松は言う。
正統的なダーウィニズムのなかにも、「発生的制約」という考え方がある(アルバーチ、1982 13)/メイナード・スミス、1985 14))。遺伝子に何らかの変異が起きても、発生上の内的な要因によってその変化が出現することが抑えられる、 という考え方である。ある変異がそのまま異なる進化系列にもちこまれ、それぞれの系列で独立にこの制約がとれて同一の形質が出現すれば、それは平行放散進化のような現象となって現れるだろう。またタイプ・スイッチング仮説は、種の多様性は遺伝子にあらかじめ構造化された多様性の現れである、とする構造主義者をも喜ばせている。
構造主義の先駆けである柴谷篤弘は、アフリカ・ビクトリア湖に棲む熱帯産の魚カワスズメの爆発的多様化も、タイプ・スイッチング的な内在的な力によるものではないか、と考える。ビクトリア湖はたかだか100万年前にで きた湖だが、そこにその土地固有のカワスズメが200種も 棲んでいる。形態はさることながら、驚くべきはその生活様式の多彩 さである。藻を食べるもの、プランクトン食のもの、あるいは、湖底の有機堆積物を食べるもの、そして貝を食べたり、昆虫を食べるもの、他の魚を食べ るもの、といった具合なのだ。この多様化が、ミトコンドリア遺伝子を使った最近の系統解析によれば、おそらく1つの祖先種からたかだか20万年の間に生じたものであることが示されている(メイヤー、ウィルソンら、エービス 1990)15)16)。このような急速な多様化を、自然選択の結果 とするのは困難なのではないか、と柴谷は言う。
この仲間も放散/平行進化?

①~④ビクトリア湖のカワスズメ(写真=東山泰之)、⑤アオバセセリ(写真=山口進)
アオバセセリの研究でよく似た平行変異の現象を指摘(1992)17)した千葉秀幸(ハワイ大、昆虫学科)も、「遺伝子にあらかじめ可能性として存在する多様性」が、何らかの理由で出現していくのが種の多様化である、と考える一人である。千葉は、大澤の言うタイプ・スイッチングがチョウの世界でも頻繁に見られると指摘する。たとえば 、アゲハチョウやタテハチョウのようなセセリチョウがいるし、東南アジアとアフリカに分布するアオバセセリ亜科の翅の紋様と、非常によく似た紋様を見せるチャマダラセセリの仲間がアメリカにいたりするのである。
だが、本当に自然選択では急速な形の変化や種の爆発 的な多様化を説明できないのだろうか。ダーウィンが生物進化を確信したガラパゴス諸島に棲むダーウィン・フィンチの最近の研究は、自然選択が驚くほど早く形態を変化させるという結果を出している。
この文鳥に似た小鳥は、くちぱしの形が食生活の仕方によってさまざまに多様化している。種子を食べるタイプはがっしりしたヒワ型、サボテンを食べるタイプは長く下方に湾曲、昆虫を食べるタイプはほっそりしている、といった具合に大きく12のグループに分かれている。米国プリンストン大学のグラント夫妻らの20年に及ぶ観察によると、自然選択の力は非常に強力で、旱ばつなどの自然災害が頻繁に起これば、わずか100年で種は別の種へと変わりうると計算される。彼らによると、遺伝子の組み換えは交雑によって起こり、環境に適応した雑種が新しい種へと進化していくという(『フィンチの嘴(くちばし)』ジョナサン・ワイナー、1994)18)。
とはいえ、このフィンチの例も、自然選択による新しい種の出現を現実に確認したわけではない。 中立的なタイプ・スイッチングによって多様なくちばしがまず生まれ、自然選択によって生き残ったのが現在の多様な種である、との可能性も否定はできない。おそらく現実の進化は、ダーウィン的な自然選択やタイプ・スイッチングなどいくつもの要因が重なり合った複合的な大ドラマである、とするのが正解なのではないか。
ミトコンドリアDNAによるオサムシ解析は、少なくとも次の事実を明らかにした。「形態におけるみかけ上の連続性は、進化の連続性を示しているとは限らない」。 また「みかけの形態が似ていても、両者の分岐時期が新しいとは限らない」。つまるところ、「これまでの種とされている区別 は必ずしも系統を反映」せず、「種とは何か」との本質的な疑問へとたどり着く。
系統の分岐と地史とが、かなり密接につながっていることも指摘された。
オオオサムシは日本の固有種だが、DNA解析によればオオオサムシは約1000万年前に放散を始めたと推定される。これは最新の地質学的研究が明らかにした、日本列島が大陸から分かれた時期とほぼ一致する。ダーウィン・フィンチの研究も、進化がガラパゴス諸島における旱ばつや洪水といった現象と密接につながっていることを示した。進化は、いまやダイナミックな地球的変動との関連でとらえられるようになっている。かつて、進化は検証できないから科学ではない、といわれたが、いまや世界各地で実証的な進化の研究が急速に進み、新しい生命観・世界観を切り開きつつあるのである。
<引用文献>
1)Zhi-Hui Su, O.Tominaga, T.S.Okada, T.Ohama, R.Ishikawa, E.Kajiwara, K.Nakamura & S.Osawa, J.Mol. Evol. submited
2)Zhi-Hui Su, T.Ohama, T.S.Okada, K.Nakamura, R.Ishikawa & S.Osawa, J.Mol.Evol. in pass 3)Andrew V.Z.Brower, Proc.Natl. Acad. Sci. USA. 91:6941-6495,1994
4)Lynn H.Throckmorton, Systematic Zoology 14: 221-236,1965
5)Terry Erwin, The Coleopterists Bulletin, 3: 74-75,1982
6)M.Kimura, The Neutral theory of molecular evolution. Cambridge University Press, Cambridge, 1983
7)T.Miyata, K.kuma, N.Iwabe & N.Nikoh, Jpn.j. Genet 69:473-480, 1994 N.Iwabe, K.kuma, T.Miyata, Mol.Biol. Evol. inpress 1996
8)Ishikawa, R.Bull.natn.Sci.Mus.16: 191-215, 1973
9)倉田貞滋・久保田耕平 昆虫と自然 30:13-19, 1995
10)David Gourson, Zoological Journal of the linnean Society, 107: 65-71,1993 11)A.H.Porter & A.M.Shapiro, Annals of the Entomological Society of America 83: 107-114,1990
12)小松正 日本昆虫学会第55回大会口頭発表 1995
13)P.Alberch,Evolution ando Deveropment, ed. J.T.Bonner, pp. 312-332,Dahlem Konferezen, 1982
14)J.Mynard Smith et al, The Quarterly Review of Biology, 60: 265-287,1985 15)A.Meyer, A.C.Willson et al, Nature 347: 550-553, 11, 1990
16)J.C.Avise, Nature 347: 512-513, 11, 1990
17)千葉秀幸 Butterfiles 第2号,39-42, 1992 18)J.Weiner, The Beak of the Frinch, Alfred A.Knopf,Inc. 1994
18)J.Weiner. The Beak of the Finch, Alfred A.Knopt, Inc. 1994















