RESEARCH
後ろ足の位置の多様性を生み出すしくみ
四肢動物の骨格を見てみると、背骨の長さや背骨を構築する脊椎骨の数は動物種によって異なるが、後ろ足(後肢)の付け根は必ず胴体の終端にあたる仙椎の領域に形成される骨盤に接続している。首長竜などの絶滅種でもこれは同じだ。今回、様々な四肢動物の発生過程を追うことで、進化の過程における後肢の位置の多様化のしくみが見えてきた。
1.四肢動物の骨格の基本構造
四肢動物には、胴体を貫く背骨(脊椎骨)がある。脊骨は形の違う脊椎骨が首の方から頚椎(けいつい)、胸椎(きょうつい)、腰椎(ようつい)、仙椎(せんつい)、尾椎(びつい)と連なったものである(図1A)。脊椎骨の長さや数は、動物種によって異なるが後ろ足(後肢)の付け根は必ず胴体の終盤にあたる仙骨(仙椎由来)を含む骨盤に接続している。これは、今生きている動物だけでなく既に絶滅してしまった恐竜や首長竜に至るまで、あらゆる動物に共通している(図1B)。つまり、脊椎骨の数や形は進化の過程で大きく変化したにもかかわらず、後肢は例外無く仙椎の位置に形成されるのである。ここで、なぜ後肢は必ず仙骨に隣接する位置にあるのかという問いが生まれる。これは四肢動物の形態形成の際にプロポーションの変化を生む後肢の位置の多様化のしくみを知ることにもつながるはずである。
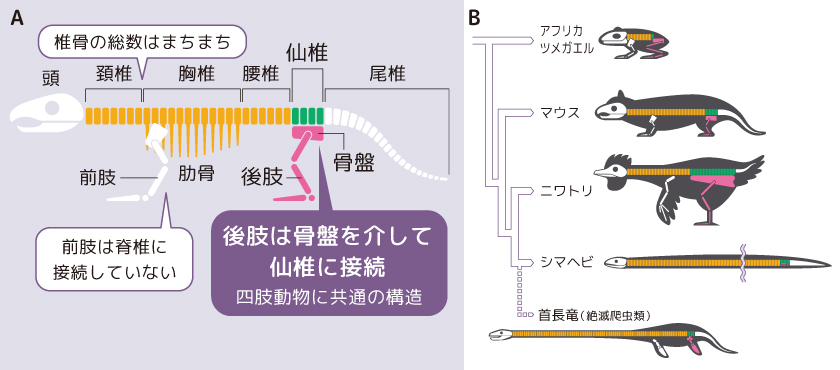
(図1) 四肢動物共通の構造
(A)四肢動物の基本骨格
(B)進化の過程を経ても基本骨格は共通
2.仙椎と後肢の発生メカニズム
四肢動物の脊椎骨は、胎児期にからだの中央を通る前体節中胚葉(PSM)に由来する体節からできる(図2)。もちろん仙椎もである。一方、後肢はPSMに隣接する側板中胚葉(LPM)の一部の細胞が増殖して生じる膨らみ、肢芽(しが)が伸長して形成される。つまり仙椎と後肢は全く別の組織から生じ、両者の細胞は混じり合うことなく独立に形成される。それにも関わらず後肢は必ず仙椎の位置にできるのである。このように別々に形成される両組織の位置を同調させるしくみは何かを見つけるべく、私たちは研究を進めた。
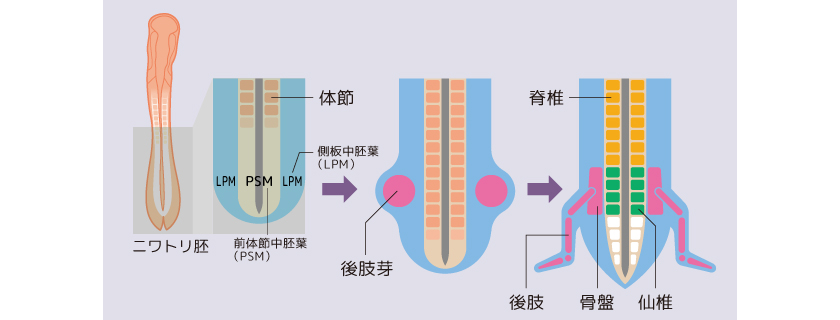
(図2) 仙椎と後肢の発生
仙椎と後肢はそれぞれ独立に発生する
3.かたちづくりを制御するGDF11
脊椎動物の初期の胚発生において、体の前後軸に沿った組織の位置は、転写因子としてはたらくホメオティック遺伝子(Hox遺伝子)の発現パターンによって大まかに決定されていることが知られている(図3)。PSMから発生する椎骨の形成もHox遺伝子によって制御されており、仙椎の形成にはHox11が重要であることが明らかになっている。実は、LPMでも、Hox遺伝子が発現しているが、後肢形成での詳細なはたらきは明らかでない。
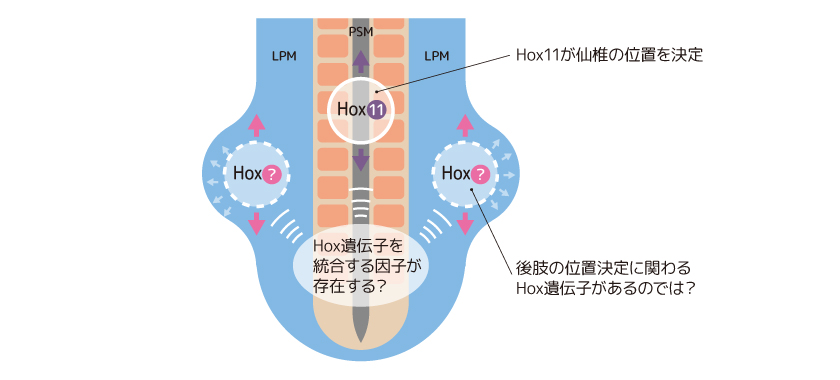
(図3) 体の前後軸を決めるHox遺伝子
調べてみると細胞の増殖分化に関わるGdf11遺伝子がはたらかないと仙椎と後肢の位置が脊椎骨数個分、体の後方にズレるというが報告があった(McPherron et al., 1999)。そこで私たちは、この遺伝子の作る分泌タンパク質であるGDF11がPSMとLPMで独立に発現するHox遺伝子の発現を上流で統合している可能性があると考えた。私たちは、体の発生過程を観察しやすいニワトリ胚を用いてGDF11の後肢の発生との関わりを追い、まずGdf11遺伝子が胎児期に脊椎骨が作られるPSMでのみ発現し(図4A)、仙椎の形成に重要なHox11の発現を誘導することを確かめた(図4B)。次に、LPMにGDF11タンパク質を添加したところ、LPMから後肢が形成された。GDF11はPSMに隣接するLPMにおける後肢の形成にも影響を与えているのである(図5A)。GDF11はさらにLPMで後肢の領域に特異的に発現するHox9-13遺伝群の発現も誘導していた(図5B)。つまり、PSMでしか発現しないGDF11が仙椎だけでなく、LPMで後肢の形成に関わるHox遺伝子の発現をも、その上流で制御しているらしいのである。
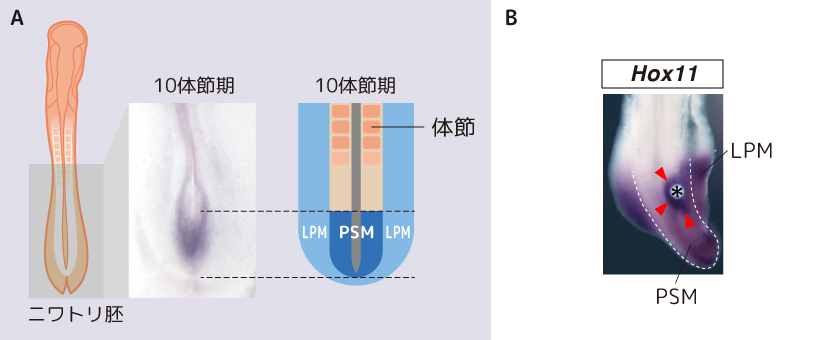
(図4) PSMにおけるGDF11のはたらき
(A)GDF11遺伝子はPSMでのみ発現する
(B)GDF11は仙椎の形成を制御するHox11の発現を誘導する。*はGDF11タンパク質を吸着させたビーズ
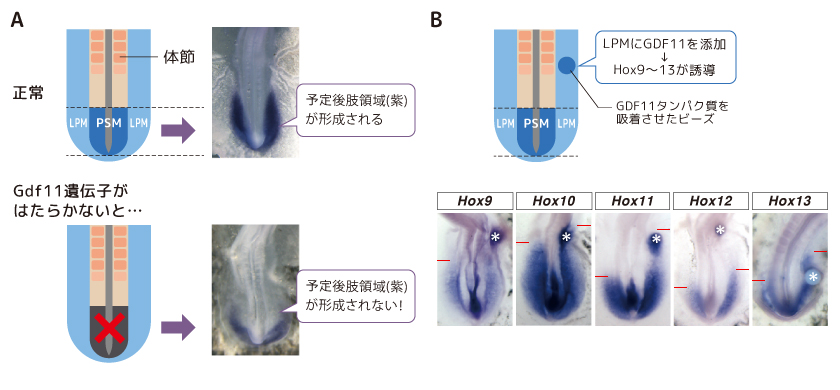
(図5) LPMにおけるGDF11のはたらき
(A)GDF11がはたらかないニワトリ胚では後肢予定胚域が形成されない。
(B)GDF11はLPMにおけるHox9-13の発現を誘導する。 *はGDF11タンパク質を吸着させたビーズ
しかし、先述したように、Gdf11遺伝子の発現はPSMでしか見られない。そこで私たちは、PSMで発現し細胞外へ分泌されたGDF11がLPMへ到達するものと仮定して、その拡散分布を確かめるためにProximity ligation assay(註1)という手法を用いて、GDF11タンパク質が結合した受容体の検出を試みた。その結果(図6)に示す通りPSMで発現した分泌タンパク質GDF11がPSMとLPMの二つの領域に作用し、仙椎と後肢の形成を誘導していることが明らかとなった。
このしくみによって、多様なプロポーションへと進化しながらどの動物も、後肢は必ず仙椎の位置に形成されるという四肢動物の形の共通性が保たれているのだ。
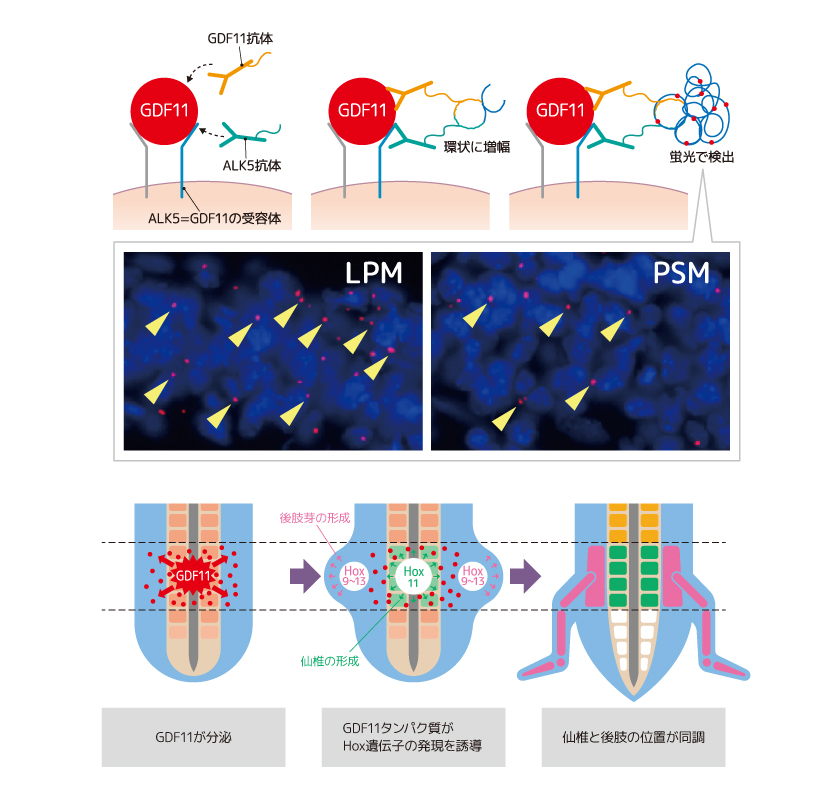
(図6) 仙椎と後肢の発生を同時に制御するGDF11
GDF11がPSMからLPMにも分泌され両者に同時に作用することで、仙椎と後肢の位置は同調する
(註1) Proximity ligation assay
オリゴヌクレオチド(DNA等)が結合した抗体を使って、細胞及び組織内のタンパク質相互作用を視覚化する方法。 この抗体の距離が近接したとき、オリゴヌクレオチドの合成・伸長反応が起こり増幅されたオリゴヌクレオチドを標識することで,相互作用を検出できる。
4.後肢の多様化とGDF11
そこで私たちは、動物種間で後肢の位置の違いが生まれるしくみも調べてみたいと考え、脊椎動物の中で胴体の短いものから、胴体の長いものまで合わせて8種の動物胚を採取し、胚発生過程でのGdf11遺伝子の発現パターンを比較した(図7)。その結果、カエルやカメなど胴体の短い(頭から後肢までが近い)ものほど発現のタイミングが早く、エミューやヘビなど胴体の長い(頭から後肢までが遠い)ものは、発現のタイミングが極めて遅いことがわかった。この結果から、四肢動物の進化に見られる後肢の位置の多様化は、発生過程におけるGdf11遺伝子のはたらくタイミングのズレによって生じると考えてよかろう。
そこで、発生過程におけるGdf11遺伝子のはたらくタイミングを変えてみた。ニワトリ胚でGDF11タンパク質が作用するタイミングを早めたところ、後肢は前肢に近い位置に形成され胴体が縮み、逆に遅らせると、後肢はより尻尾に近い位置に形成され胴体が伸びた。こうして発生過程におけるGDF11タンパク質のはたらくタイミングの重要性が証明できた。
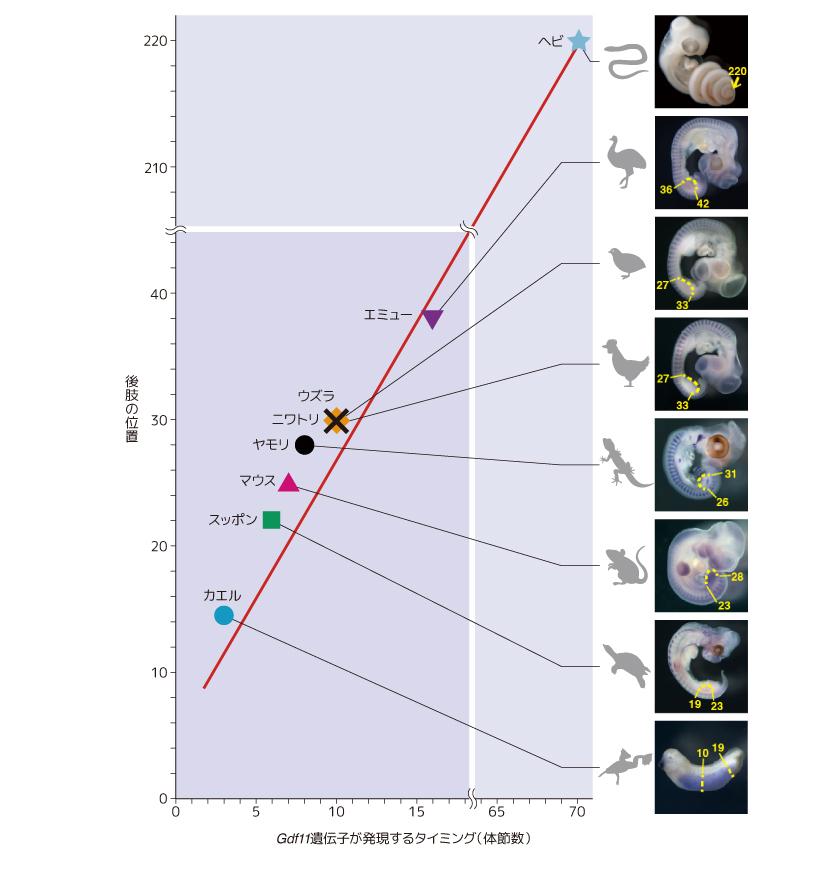
(図7) 8種の生物種におけるGdf11遺伝子の発現時期
Gdf11遺伝子の発現するタイミングが後肢の位置を決める
5.まとめと今後の期待
Gdf11遺伝子はヒトを含むすべての脊椎動物に存在する。地球上の様々な形態をもつ脊椎動物のほとんどの種がその発生過程において、これまで語ってきたしくみをはたらかせているはずである。Gdf11遺伝子というたった一つの遺伝子がはたらくタイミングのズレが形態の多様な表現につながっているのである。生物の大進化は、体の形を作る多くのHox遺伝子の変化によって引き起こされたと考えられてきたが、今回の研究で、GDF11が複数のHox遺伝子のはたらく場所を上流で制御していることがわかった。このため、意外と少数の遺伝子の変化によって生物の形態の大進化がもたらされた可能性が考えられる。今後、胎児期におけるGDF11の作用をさらに詳細に調べ、下半身全体の器官の位置を決める発生のしくみを知りたいと考えている。

鈴木 孝幸(すずき・たかゆき)
1999年慶應義塾大学理工学部応用化学科卒業、2004年 奈良先端科学技術大学院大学バイオサイエンス研究科後期博士課程修了。博士(バイオサイエンス)。日本学術振興会特別研究員、海外特別研究員(於ウィスコンシン大学)、さきがけ研究員(兼任)などを経て、2018年より名古屋大学大学院生命農学研究科准教授














