RESEARCH
トマトの実を育む細胞壁の変化
花は受粉が終わると、次世代を生む種子を育む実をつくる。この過程で、役目を終えた花は「落花」により、熟した果実は「落果」によって茎を離れる。これは劣化や衰えによる現象のように見えるが、実はここに植物細胞の巧みなしくみがあることをトマトで明らかにした。
CHAPTER
1.植物の戦略
花は受粉が終わると、次世代を生む種子を育む実をつくる。この過程で役目を終えた花は「落花」、熟した果実は「落果」によって枝を離れる「器官脱離」が見られる。このとき脱離する境界となるのが「離層」である。植物は発生の過程で、幹と葉や花などの器官をつなぐ枝(葉柄、花柄)に離層をつくることがわかってきた。離層部は、特殊に分化した小さな分裂組織細胞が軸を横切っており、繊維が存在しないので、力に弱い構造になっている。落花、落果などの器官脱離は、老化による衰えに見えるが、実は植物に本来備わったしくみなのである。植物細胞に特有の細胞壁に注目し、そこからトマトの開花や結実に伴う器官脱離を調べたところ、それが植物の生存戦略の一つと考えられることがわかった。
例えばトマトの花での落花を見ると、触ったか触らないかわからないくらいの強さで触れただけでも落花するものが多いが、なかには触っても落ちないものもある。両者の違いに注目したところ、受粉しなかったものが落ちやすいことがわかった。花は生殖のため、そして種子をつくるためにあるわけで、受粉が成功しなかった花は、植物にとって栄養を使う負荷となるので落ちやすくしているのだ。不要な花は取り除き、養分を種子形成に分配する生存戦略である。
一方、熟した果実に見られる落果は、触った程度では落ちない。果実は種子の成熟の場であり、動物の食物となり種子を散布する役割をもっている。トマトの場合、果実を食べ種子を運ぶのは主に鳥類である。熟した果実の中には発芽率の高い種子が含まれており、甘く栄養価の高い熟れた果実を鳥は喜んで食べる。果実を提供することで、広い範囲を動き回れる鳥に種子を遠くに運んでもらい生活域を広げるのが植物の戦略なのだ。プチトマトでは、鳥が枝に生ったままの果実をついばむこともあるが、容易にもぎ取る様子もみられる。運ばれやすいように簡単な刺激で切り取られるしくみをつくっているのである。
トマトの小花柄の離層では受粉時の花、成熟時の果実それぞれで異なる二段階の器官脱離のしくみがあることがわかっている(図1)。
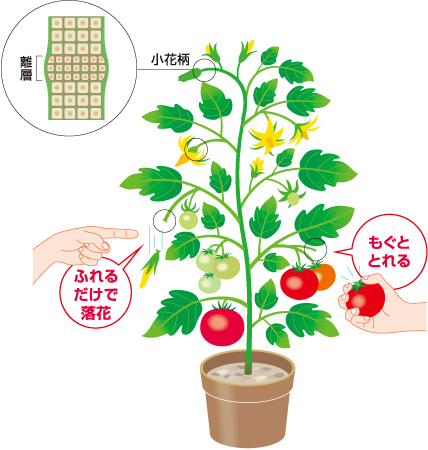
(図1) トマトの落花と落果
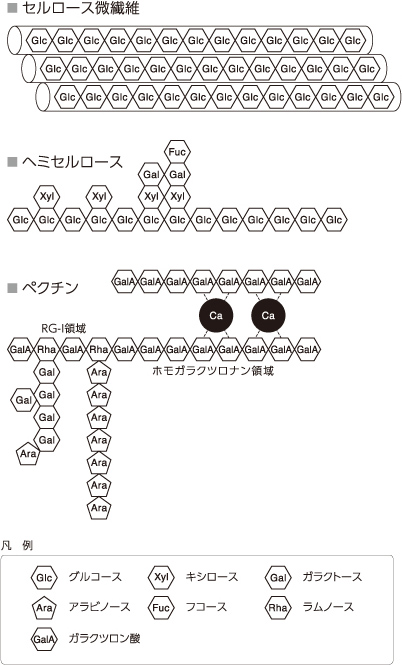
ここで触っただけで落ちてしまう受粉に失敗した花の離層と十分に成熟して鳥に運んでもらうためにもぎ取りやすくなった果実の離層とでは、どのような違いがおきているのかという問いが生まれる。私たちは、花の受粉前後と果実の成熟段階、それぞれの離層の細胞の細胞壁を構成する多糖に注目した。細胞壁を構成する主な成分は、セルロース、ヘミセルロース、ペクチンの多糖であるが(図2、註1)、これらは構造を支えるだけではなく、細胞の性質を表すと考えられている。そこで、これらの多糖を組織染色および免疫抗体染色を用いて可視化し、顕微鏡下でその疑問を解こうと考えた。
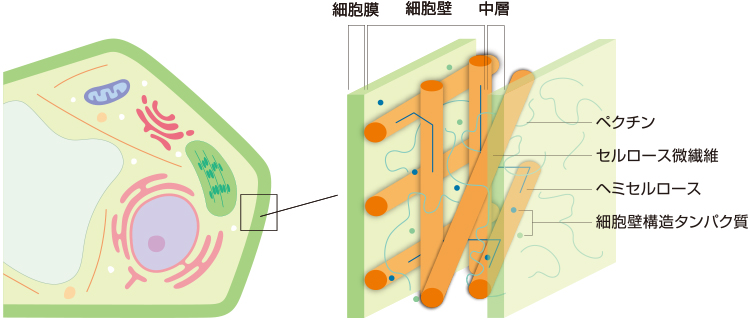
(図2) 細胞壁の構造モデル
(註1)
2.落花のしくみ
落花のときの離層の様子を調べると、離層を形成する細かい細胞層は脱離する花の側ではなく茎の基部側に残り、切り口を覆うキャップの役目をする。このキャップ構造の細胞壁を構成する多糖の組成を調べたところ、ヘミセルロースの組成が変化し、キシログルカンとアラビノガラクタンが蓄積していることがわかった(図3)。
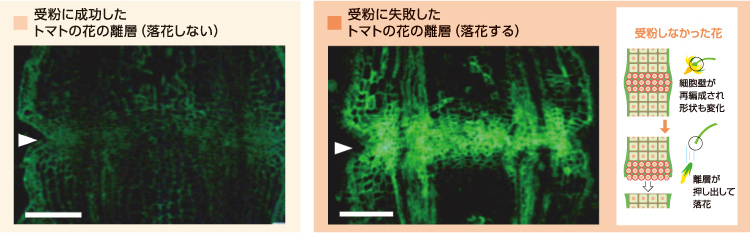
(図3) 落花における離層の細胞壁構造の再構築
受粉に失敗し落下するトマトの花の離層では、脱離する前にキシログルカンの蓄積が起きる。
この細胞壁の再編成は、受粉して維持される花では見られず、落とされる花でだけおきていた。この変化を捉えるため、多糖の再編成に関わる2つのタンパク質に注目した。一つは、エンド型キシログルカン転移酵素/加水分解酵素(XTH)であり、キシログルカンの糖鎖の切断やつなぎ換え反応を触媒し、細胞壁の構築や再編過程で中心的な役割を担う酵素である。もう一つのエクスパンシンは、セルロースと他の多糖との架橋を離し、細胞壁構造のゆるみに直接的に関わっている。受粉しなかった花では、XTHは開花1日後の離層で最大量を示しており、早い時期にキシログルカンの糖鎖を操作し、その後の細胞壁の再編成に関わる反応を促進していると考えられる。また、エクスパンシンは、脱離がおこる時期の離層で局在量が最大となり、多糖の架橋の変化が脱離に積極的に関わることを示した。そこで、離層の細胞では、キシログルカンの変化とエクスパンシンのはたらきで細胞の形が球状となり、押し出すような形で脱離が生じていると考えてよかろう。実を結ばない花では、この細胞壁の再編成によってつくられるキャップ構造とエクスパンシンによる細胞の変形によって落花をうながしているのである。
3.落果のしくみ
落果のしくみは、落花とどのように違うのであろうか。受粉した花の離層は果実の成熟までは丈夫に保たれ、実が熟れて種が育つとうまくもぎとられるキャップ構造ができる。このキャップ構造の組織を染色すると、細胞壁に木質を構成するリグニンが蓄積していた(図4)。
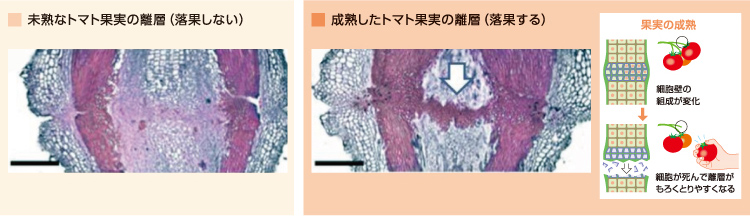
(図4) 落果における離層の細胞壁構造の再構築
成熟して落果するトマトの花の離層では、脱離する前にリグニンの蓄積が起きる。
リグニンの蓄積には、二つの可能性が考えられる。一つ目は、道管のように物理的強度を増した二次細胞壁の形成であり、二つ目は細胞が死んでいるだけというものである。二次細胞壁は強固な細胞壁であり、新たな形成にはセルロースの合成を伴うことが知られている。そこで、落果過程の離層においてセルロース合成がおきているかどうかを調べたが、セルロース合成酵素は増加しておらず、二次細胞壁の形成ではないことがわかった。また、落花で見られたエクスパンシン、XTHともにこの場合はほとんど蓄積がない。そこで果実の離層では、果実の成熟の過程で細胞が死んでいくと考えている。リグニンをもつ細胞が死ぬと空洞化してもろい構造になり、もぎとりやすくなる。果実では積極的に落とすのではなく、運搬者が訪れるまで待って取らせる戦略をとっているのである。
花と果実共に、器官脱離の時に変化するのは、花柄の離層の周辺という極めて微細な細胞層であった。このように極めて限定された細胞層でおこる現象であったためにその構造もはたらき方もわかっていなかったのである。離層では、花でも果実でも同じようにキャップ構造をつくり、落ちる準備をするのだが、花では細胞壁の組成や構造を変えて細胞を変形して離れやすくし、果実では死んだ組織となり外からの力に任せる方法を取っている。同じ組織を構成する細胞壁を花と果実では異なるつくり変えをし、繁殖の目的を果たす植物の生存戦略が、この小さな細胞層から見えてきた。
4.果実が熟れるしくみ
果実には、種子を散布する動物にとって魅力的な食物となるための戦略がある。エネルギー源となる炭水化物、ビタミン類等を蓄積し、容易に食べられるよう熟して軟化するのだ。そのため、果実の軟化と十分な栄養分の保持を両立するしくみが必要である。果実の軟化は、主な可食部である果皮の細胞壁、特に細胞壁成分中に約50%と豊富に含まれるペクチンの分解による低分子量化であると考えられていた。しかし、ペクチンの分解を抑えても果実の軟化を止めることはできないことがわかり、細胞壁のペクチンの分解は直接の原因ではないと考えざるを得ない。そもそも分解だけであれば、軟化して成熟した赤い果実は、硬い未成熟な緑の果実の分解物でしかないことになる。しかし実際は、果実の成熟は熟した果実へのつくりかえであり、その際果実専用の「しなやかな細胞壁」へと再構築がおきているのである(図5)。
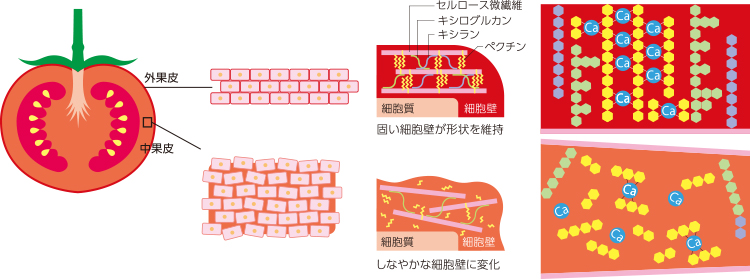
(図5) トマト果実の軟化のしくみ
外果皮では、ペクチンがカルシウムで架橋され、ヘミセルロース(キシラン、キシログルカン)が合成される。
中果皮では、低分子のペクチンが増加し、ヘミセルロースのつなぎ換えがおきている。
細胞壁の組成を調べたところ、柔らかな中果皮組織では、キシログルカンがつくられ、分子量の小さいペクチンが増えていた。さらに、柔らかく液化した中果皮組織を包む外側の外果皮や内側の皮は、キシランを蓄積し、筋張った組織に多いより硬い細胞壁へと再編成していた。熟する過程で果実は単に分解するだけではなく、細胞壁での組織ごとに異なる合成がおきているのである。また皮では、カルシウムとペクチンの架橋が非常に多いことも示された。このことからカルシウムがトマト果実の細胞壁の再構成に重要であると考えている。
トマトの落花や落果、そして果実の軟化を見てきたが、細胞壁は、ただ単に壁として存在しているのではなく、植物の生存戦略の中で極めて柔軟な表現をしていることがわかった。離層の細胞壁を変化させることにより脱離しやすいような組織を形成したり、果実の軟化の際には、ペクチン量を増やし柔らかい細胞壁へと変化することで組織を柔らかくしたり、植物の生存戦略の切り札とも言える役割を担っているのである。今後、細胞壁がどのように制御されているかがさらに明らかになれば、より深く植物の成長、発達が理解できるようになっていくと期待している。トマトでの結果をもとに、イネの細胞壁再編成の解明も行いたいと考えている。

岩井宏暁(いわい ひろあき)
筑波大学生物学類卒。同大学院生物科学研究科修了。博士(理学)。日本学術振興会特別研究員を経て、2003年筑波大学講師。2014年より同大学生命環境系植物生理学研究室准教授。