RESEARCH
葉緑体と植物進化の光と陰
シアノバクテリアの共生によって葉緑体を獲得した真核生物が植物である。葉緑体は、それまで光に依存せず生活していた宿主の生き方を根本的に変えた。異なる生きものが1つの細胞として生きはじめた時、何が起きたのだろう。
1.植物をつくったオルガネラ
植物の葉を顕微鏡で観察すると、楕円形で緑色の葉緑体を見つけることができる。葉緑体は植物が進化の過程で獲得した光合成を行なう細胞内小器官(オルガネラ)だが、本来は酸素発生型細菌であるシアノバクテリアが共生したものである。その結果、宿主の真核生物は光をエネルギー源として水と二酸化炭素から糖をつくる独立栄養性を獲得し、それまでの生き方が根本的に変わった(図1)。
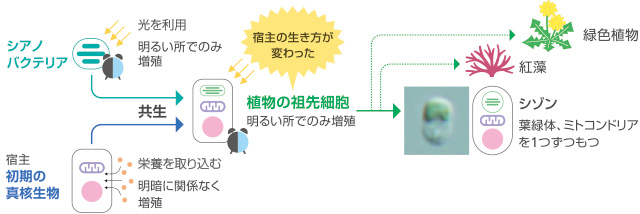
(図1) シアノバクテリアと真核生物の共生による植物の誕生
植物は基本的に暗所で増殖、成長しないが、葉緑体を獲得する前の宿主は外から栄養を取り込む従属栄養性であり、光に依存せず増殖していたはずである。進化の過程で宿主と、外から入り込んだ生物が変化した光依存性の葉緑体とが連携し共生のしくみができたのである。共生の成り立ちを知りたいと、私たちは単純な構造をもつ単細胞紅藻、シゾン(Cyanidioschyzon merolae)に注目した。
2.シゾンが物語る共生のはじまり
シゾンは、ミトコンドリアと葉緑体を1つずつもち、その他のオルガネラも最小単位から成る単純な構造をもつ。環境変動の少ない硫酸酸性の温泉の中で暮らしており、植物が誕生した当時の原始的な姿を留めていると考えられる。私たちはシゾンを用い、葉緑体から核へと情報を伝えるしくみの研究を行なった。光を受けると、葉緑体では光合成をはじめとする様々な活動が活性化される。植物細胞としては、この葉緑体の状態が核に伝わることで、必要に応じて核遺伝子をオン・オフするしくみを必要とするはずだ。シロイヌナズナなど高等植物での研究から、葉緑体でつくられるクロロフィル(葉緑素)合成中間体が細胞質に放出され、核遺伝子発現を制御する信号として働くことが知られている。そこで私たちは、シゾンも同様のしくみをもつことを予想し、暗所で増殖の停止したシゾン細胞に複数のクロロフィル合成中間体を作用させ、核遺伝子のオン・オフを調べた。その結果として、中間体の1つMgプロトポルフィリン IX(Mg Proto)が、遺伝子の転写でなく、核ゲノムの複製を開始させるという予想外のことが見つかったのである。
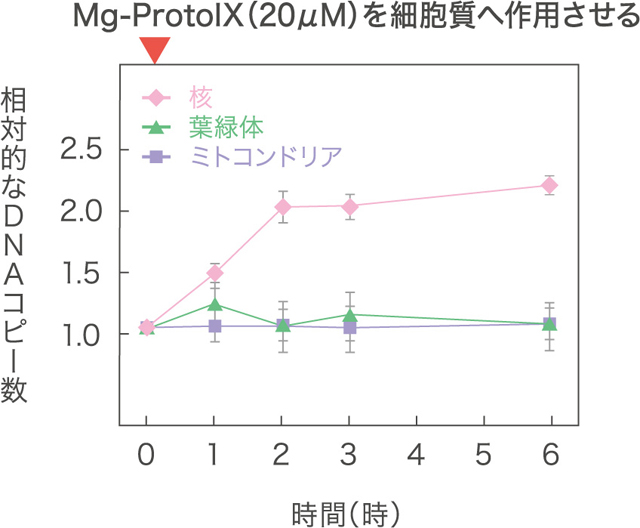
(図2) Mg Protoの作用による核・オルガネラDNAコピー数の変化
Kobayashi et al., Nature Cell Biol. 13, 483-487.(2011)より改変。
3.葉緑体からの合図ではじまる核の複製
暗所でのシゾンの細胞周期は、DNA複製をするS期の前段階、G1期で停止している。この時の細胞内の様子を調べると、S期の開始に必要なサイクリン1がタンパク質分解酵素複合体、プロテアソームにより分解されていた。この時、Fbx3というタンパク質が、分解されるための目印(ユビキチン)をサイクリン1に付けていることがわかった(図3)。
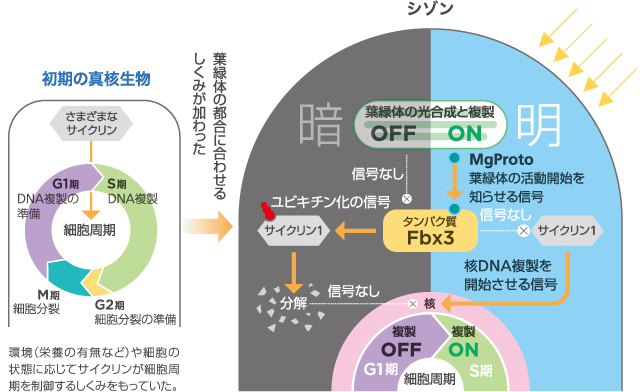
(図3) シゾンの葉緑体と核ゲノム複製が連動するしくみ
一方明所では、葉緑体が活性化してMg Protoが細胞質へ放出され、Fbx3と結合してユビキチン化を防ぐ。このはたらきによって、明所ではサイクリン1の分解が起きず、その結果、S期への移行が始まり核ゲノム複製が開始するのである。今回見出した、明か暗かという光環境の変化と連動する細胞周期の制御は、シアノバクテリアが共生する前の真核細胞がもっていた細胞周期の調節システムに「暗所で開始を阻害し、明所でその阻害を解除する」しくみがつけ加わったものと考えられる。
シアノバクテリアと真核生物が共生した初期段階を想像してみよう。おそらく、暗所では宿主は増殖できても共生体は増殖できなかっただろう。もし、宿主の細胞周期を制御するしくみが変わらなければ、共生体が増殖できないままに宿主細胞が分裂を繰り返し、結局は共生体がいない細胞が大多数になってしまうはずである。共生関係を維持するには、暗所で宿主細胞の増殖を止めるしくみが不可欠であり、これが植物が暗所で生えない大本の理由となった。私たちの研究は、このしくみがたった一つのタンパク質(シゾンの場合はFbx3)の獲得によって、可能となることを示す興味深い成果である。
4.光との関わり合いから見える植物進化の原動力
明所での活動を基本として、暗所でその機能を抑制するというしくみは、双子葉植物の発芽時にも見られる。種子が発芽する時、明所では双葉の開いた芽生えとなり、暗所で「もやし」となるのはご存知の方も多いだろう(図4)。これは、深い土中で発芽した場合、双葉を開かずに胚軸が伸びる「もやし」となることで、無事に地上まで到達するためのしくみである。暗所でも明所のように双葉を開く変異体が多数見つかっていることから、種子発芽は明所での子葉づくりが基本で、暗所でそれを抑制するしくみが後から進化したと考えられる。このような例も合わせて考えると、光合成で生きる植物にとっては光合成可能な明条件が基本であり、暗条件を生きるしくみの獲得を原動力に植物が進化してきたと言えるだろう。
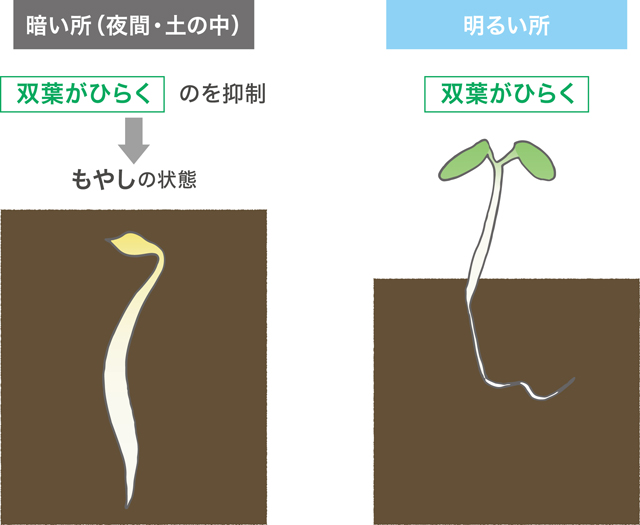
(図4) 双子葉植物の明暗での子葉づくりの切り替え
これからも、共生が成立した原始の姿を留めていると考えられるシゾンに注目し、真核細胞の細胞制御の基本的なしくみを明らかにしていきたい。そして、そこから多細胞生物も含めた多様な真核生物の生きるしくみに迫りたいと考えている。
ところで、なぜオルガネラゲノムから核へと一方向に遺伝子が移行するのだろう? また、すべての遺伝子が核へ移らずオルガネラゲノムが今も存在しているのはなぜだろう? 今のところ、遺伝子を抜き取って核がオルガネラを制御できるようになることが真核生物の生存に有利だったからとする説が有力である。オルガネラへの輸送が困難なタンパク質や、素早い発現応答が必要なタンパク質の遺伝子だけがオルガネラゲノムに今も残されているというのだ。一方、ミトコンドリア、葉緑体ともに膜を用いた電子伝達系があり、そこで発生する活性酸素で遺伝子が損傷を受けやすいため、より安全な核内に遺伝情報を退避させているという説明もされている。タンパク質を輸送する手間がかかっても、遺伝情報の損傷を避けるメリットの方が大きいという考え方である。
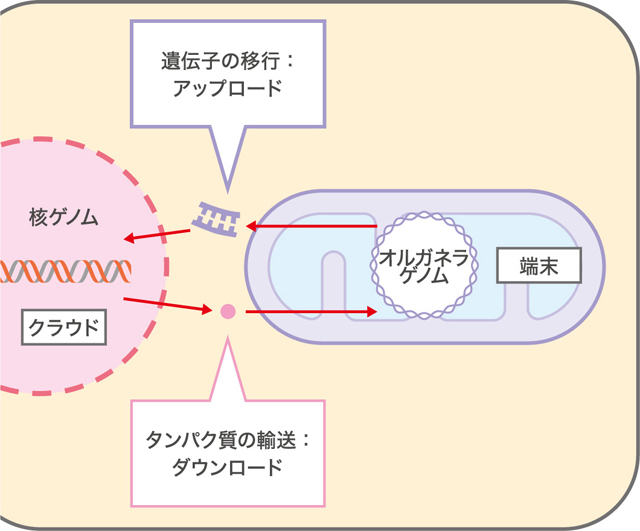
最近、私はこのようなオルガネラゲノムと核ゲノムとの共進化が、近年のコンピューターネットワーク環境の急速な進歩に似ていると考えるようになった。パソコンが普及し始めた頃には、全ての情報処理をパソコンの内蔵メモリを使って行っていた。インターネットが現在のように発展するとは想像もできず、コンピューターをネットワークに繋ぐのは簡単なことではなかったのである。今では重要な情報はパソコンの中だけでなく、ネットワークを介してクラウドサーバーに保存・共有されるシステムが当たり前になりつつある。パソコンだけでなく、スマートフォンなど多様な端末から同じプラットフォームで情報が一元管理できることは非常に便利であり、実際に使ってみてその威力を実感する。真正細菌が細胞内共生をしてオルガネラとなった当初、オルガネラの遺伝情報は全てオルガネラゲノムにコードされ、オルガネラ内で閉じた情報発現系が機能していたはずだ。核とオルガネラの間で情報を伝えるネットワークが深化するにつれ、上位のクラウドサーバーに相当する核ゲノムに遺伝子が移行(アップロード)していったと捉えてみよう。端末にあたるオルガネラは、安全な核に情報を預けたために、日々タンパク質(情報)をダウンロードしながら生きている訳である。以前はパソコンのメモリがクラッシュする度に、何度も悲しい思いをさせられた。クラウドサーバーの恩恵を受ける私には、なぜ核ゲノムに情報が集約されていったかがわかる気がするのである。

田中 寛(たなか かん)
1990年東京大学大学院農学系研究科博士課程修了。農学博士。東京大学分子細胞生物学研究所助教授、千葉大学大学院園芸学研究科教授などを経て、2011年より東京工業大学資源化学研究所教授。