RESEARCH
ウイルスに寄生する「小さな核酸」
細胞に寄生するウイルスに寄生するものがいるのだから驚く。サテライトと呼ばれる仲間だ。私は、植物に感染するキュウリモザイクウイルス(CMV)に寄生し、植物の症状を劇的に変えるサテライトRNAのはたらきを30年にわたり探ってきた。
1.植物に寄生するウイルス、ウイルスに寄生するサテライト
私たちはインフルエンザなどさまざまなウイルス感染症にかかるが、植物もウイルスで病気になる。キュウリモザイクウイルス(CMV)は、アブラムシによって媒介される直径30nmの正二十面体の粒子で、さまざまな植物に感染して葉に斑紋を作ったり、枯らしたりする。そのゲノムは1本鎖RNAで、自身の増殖のための複製酵素や、細胞間を移動するための移行タンパク質をコードしている。
CMV粒子には、CMVゲノム以外のものが入っていることがある(図1)。CMVに寄生しているサテライトRNAだ。その塩基配列は330〜400塩基と短く、タンパク質は作らない。サテライトRNA はCMVと共に移動し、CMVの複製酵素を使って増えるが、その塩基配列にCMVゲノムとの相同性はなく、起源はなぞである。
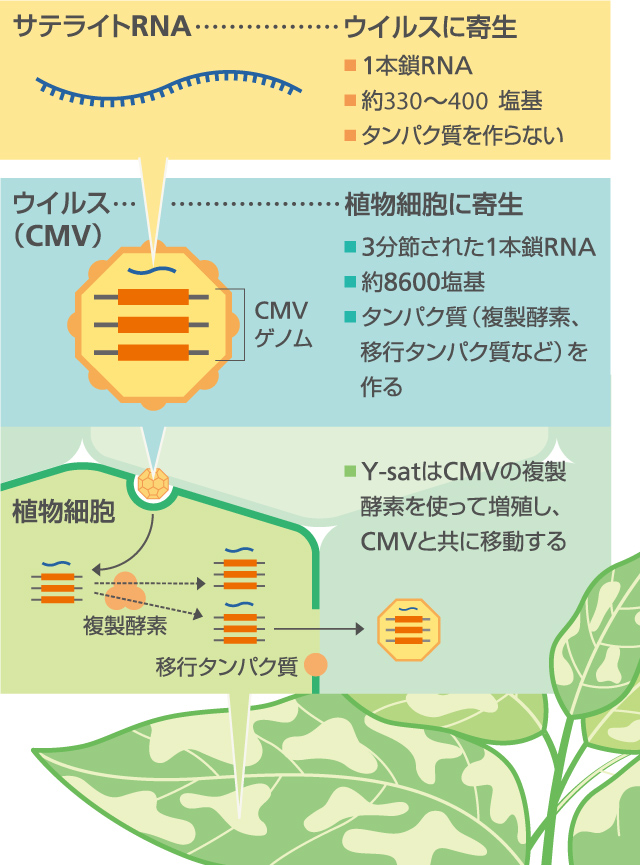
(図1) ウイルスに寄生するサテライトRNA
2.サテライトRNAと植物の関係
CMVに寄生するサテライトRNAの多くは善玉サテライトRNAと呼ばれており、CMVの防除にも利用されている。それらがCMVの複製酵素を利用することでその増殖を抑え、植物の病状を軽くしているからである。一方、症状を悪化させる悪玉サテライトRNAもおり、その1つが私たちが注目しているY-satだ。Y-satが寄生したCMVが感染すると、タバコの葉は黄色に変化する(図2)。タンパク質を作らない小さなRNAに、なぜこんなにも大きな力があるのだろうか。

(図2) Y-satがタバコ(Nicotiana benthamiana)に及ぼす影響
3.Y-sat研究の幕開けと15年の空白、そして転機
Y-satは専売公社(現JT)の高浪洋一によって、1981年に四国・小豆島のCMV感染タバコから発見された。不思議なことに、Y-satと同様の性質を備えるサテライトRNAはそれ以後もその地域でしか見つかっていない。私たちを含む複数のグループが、葉を黄化させるY-satの塩基配列領域(SYR)を明らかにしたが、その分子メカニズムは不明のままであった。その後、私たちのグループがさまざまな解析を試みたが、大きな手がかりがつかめず苦難の15年を過ごした。
転機が訪れたのは2004年。植物のウイルス防御機構「RNAサイレンシング」を抑えると、Y-satを接種しても葉が黄化しないことが、オーストラリアのグループとの共同研究でわかったのだ。RNAサイレンシングは、ウイルスやサテライトRNAの複製過程でできる2本鎖RNAを、植物タンパク質DCLが切断することで反応が始まる(図3)。最終的に、切断されたこのRNAをガイドにして、別の植物タンパク質AGO1が相補的な配列(註1)を切断する。特定のRNA配列を繰り返し分解するこのしくみは、植物にとっての免疫機構といえるが、これがY-satによる葉の黄化とも関わっている可能性が見えてきたのだ。
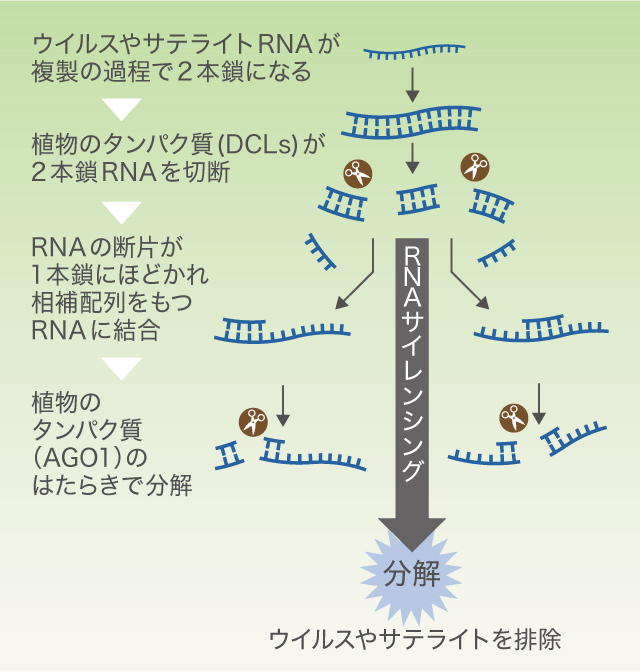
(図3) 植物のRNAサイレンシングのしくみ
(註1) 相補的な配列
RNAまたはDNAにおいて塩基対を形成する配列のこと。RNAを構成する4種類の塩基、アデニン(A)、ウラシル(U)シトシン(C)、グアニン(G)は「AとU」、「GとC」、「GとU」の組み合わせでそれぞれ水素結合による塩基対を形成する。
4.見えてきたY-satによる葉の黄化のしくみ
Y-satのみが葉の黄化を起こしていることを証明するため、CMVを感染させずに、Y-satの2本鎖RNAのみを発現する形質転換体タバコを作ったところ、確かに葉が黄化した(図4)。さらに驚いたことに、この形質転換体ではクロロフィルの合成に関わる重要な酵素遺伝子ChlIの発現が減少しており、この酵素のmRNAには、Y-satのSYRと相補性をもつ配列(YR)が存在していた(図5)。

(図4) 形質転換体をつかった実験結果
CMVに感染していなくてもY-satの2本鎖RNAが存在すれば、葉の黄化が起こる。
Shimura et al.PLoS Pathogens 7(5): e1002021(2011)より改変。
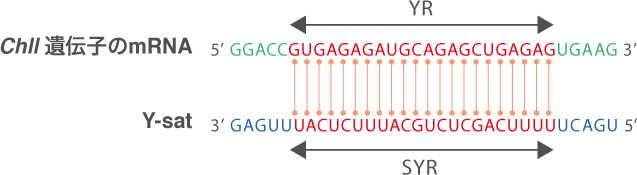
(図5) Y-satとChlIの相補配列
Y-satとChlIのmRNAの間には22塩基の連続した相補配列が存在する。
つまり、ChlIのmRNAがYR領域をもっているために、Y-satと間違えられてRNAサイレンシングによって分解され、その結果、緑の色素であるクロロフィルが作られず葉が黄化していたのである(図6)。
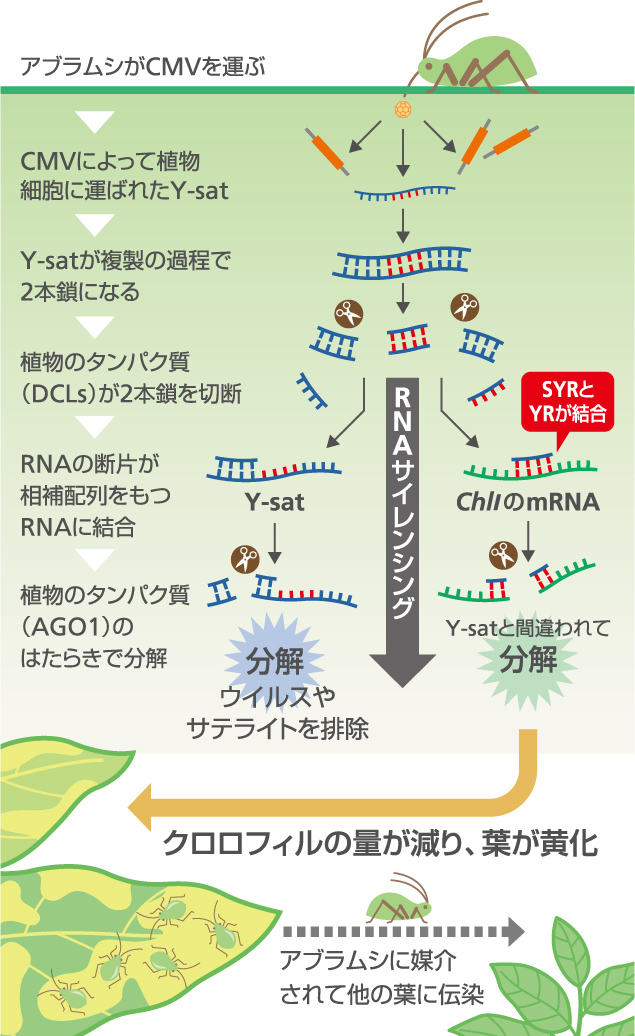
(図6) Y-satによる葉の黄化のしくみ
Y-satの研究を開始してから30年、ようやくY−satによる葉の黄化のしくみを解明するに至った。実は偶然、私たちとは独立に同じ遺伝子に同時期にたどり着いた研究グループがあり、その論文は私たちと隣り合わせで同じ雑誌に掲載されることになった。皮肉なことに、これによって私たちの仮説の信憑性は確実なものとなった。
5.そもそもサテライトRNAはどこから来たのだろうか?
サテライトRNAの起源は長年の謎で、現在も解明されていない。サテライトRNAとCMVゲノムとの間には全く塩基配列の相同性がないので、CMVが感染した細胞内でサテライトRNAが新たに作られたとしか考えられない。私たちは、Y-satのRNAの2次構造(註2)が葉緑体tRNAと相補的であることを見出しており、Y-satの起源が葉緑体にある可能性も考えている。
タンパク質を作らないサテライトRNAの存続は、複製酵素によって短時間でいかに多く複製されるかにかかっている。Y-satは、他のサテライトRNAと比べて複製能力が劣るため、本来ならば駆逐されても不思議ではないのだが、日本の一部地域で感染を広げていた。これはCMVを媒介するアブラムシと関係があるのではないかと私は考えている。実は、アブラムシは黄色を好むのだ。今回の結果から、Y-satがアブラムシに運搬してもらえるよう、アブラムシが好きな色をタバコに誘導したという解釈が引き出せる。宿主、ウイルス、媒介昆虫が関わり合う進化の中で、サテライトRNAという存在の意味を考えると新しい側面が見えてきそうだ。
(註2) RNAの2次構造
RNAが選択的に塩基同士で水素結合することにより形成する折り畳み構造。

増田 税(ますた ちから)
1981年北海道大学農学部卒業。同年、日本専売公社入社。1984~86年米国パデュー大学大学院へ留学。農学博士。1996年より北海道大学農学部助教授、現在、同大学大学院農学研究院教授。