RESEARCH
形の多様性はお母さんのお腹の中で
標本から発生、そして進化を知る
地球上に哺乳類は約6000 種、陸・海・空・地中などさまざまな場所で暮らし、それぞれの生活に適した実に多様な姿をしています。小薮大輔さんは形態の多様性を生むしくみを知るため、世界中のフィールドと博物館を訪ね、102 種もの哺乳類の胎子標本を集めその発生過程を比較しました。胎子の観察は難しいため、発生過程がわかっている種はごく一部でしたが、博物館の標本の活用で網羅的な比較が初めて可能になりました。頭部に注目すると、体重に対する脳重量の比が大きい種ほど脳を覆う骨を形成する時期が早いことがわかりました。ここから進化も語れそうです。
1.多様な形を生むしくみとは?
哺乳類の仲間は約6000種、地球上の陸・海・空・地中とさまざまな場所で暮らし、それぞれの生活に適した多様な姿をしている(図1)。この形態の多様化には、進化の過程で、胎子の成長の仕方が変化したことが関わっていると考えられている(図2)。なかでも、発生の各段階が起きるタイミングや、発生速度の変化が形態の違いを生む大きな要因になるはずだと予測されてきた。
しかし、胎子の成長は母親のお腹の中で進むために観察が難しく、哺乳類ではこれまで実験動物や家畜動物などの限られた種の発生過程しかわかっていなかった。そこで、私は博物館の胎子標本に注目し、多くの種の発生過程を比較することで、形態の多様性が生み出されるしくみを知ろうと考えた。
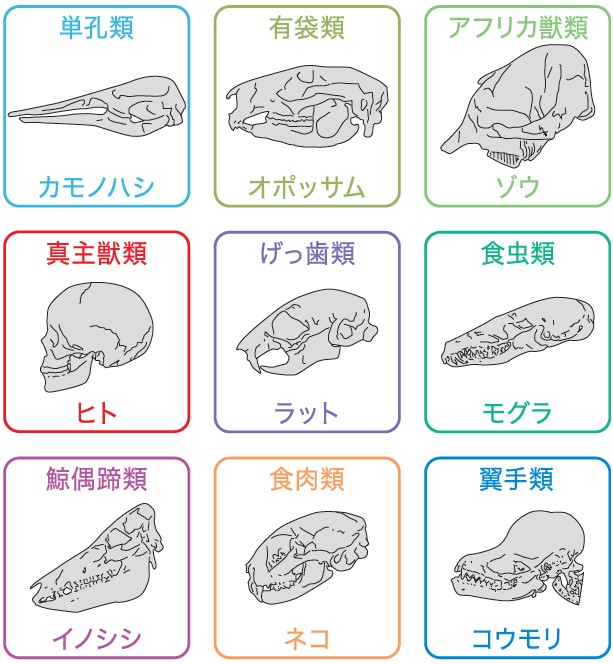
(図1) 哺乳類の頭蓋骨の多様性
母親の体内から生まれ出てきた時には、すでにそれぞれ多様な形をしており、どこが共通でどこが変化した部分なのか比較することは難しい。
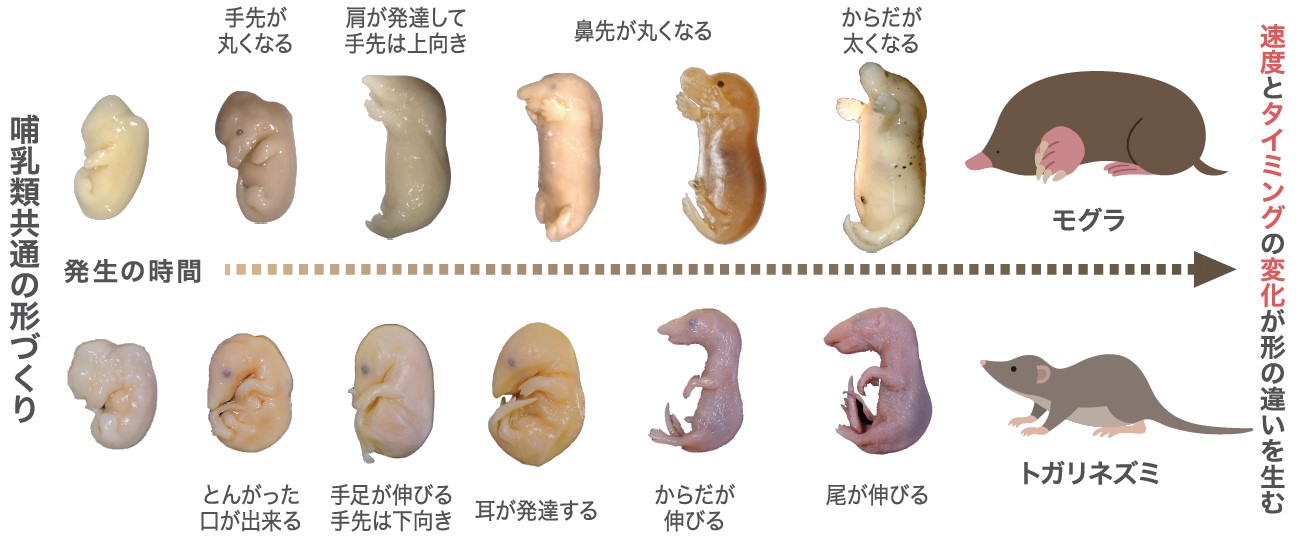
(図2) モグラとトガリネズミの発生過程
2.埃にうもれていた標本に光を
世界の博物館には100年以上にもおよぶ学術標本の蓄積が存在する。私は、2009年から世界中の博物館を訪ね胎子標本の収集を始めた。哺乳類が産む子の数は他の動物に比べて少なく、また骨格標本に比べて管理が煩雑な液浸での保存が必要になることなどから胎子標本は希少である。収集を始めた当初はどれほどの個体数が各国に保存されているかがわからなかったが、各国博物館の収蔵庫で5年間に渡って地道に収集を続けた(図3)。探せば探すほどに案外掘り出せることが分かりフィールドでの収集も加え代表的な哺乳類を網羅する102種2,000個体の胎子標本を集めることができた。こうして発生過程の網羅的な比較が初めて可能となったのである。

(図3)博物館の収蔵庫での地道な標本収集
まず、見る・聞く・食べる・呼吸するなどの重要な機能を果たす頭部の形態に注目した。哺乳類の頭蓋骨は20個ほどの骨片で構成されており、胎子の発生に伴ってそれが一つずつ形成されてゆく(図4)。形成過程のどこに種ごとの違いがあり、それが進化的にどのような意味をもつのだろう。このような問いを立て、標本での比較分析を行なったところ、脳の大きさの進化と頭蓋骨形成との関わりが見えてきた。
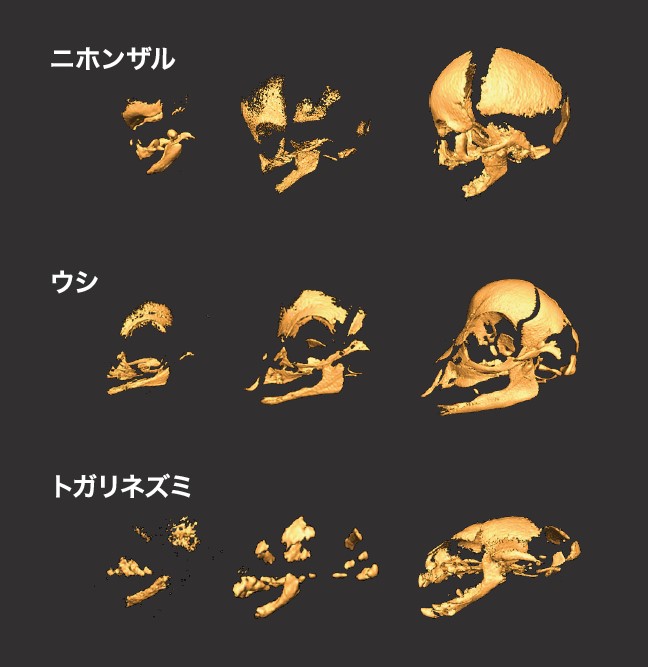
(図4) ニホンザル、ウシ、トガリネズミの頭蓋骨の形成過程
個体数が少ない種の標本は保存状態を変えないよう、最新のCT技術で画像解析を行った。
3.哺乳類の頭蓋骨形成の共通点
化石の研究から哺乳類は約2億年前に誕生し、夜行性であっために鋭い嗅覚、聴覚を備え、それまでの脊椎動物に比べて大きな脳を獲得したと考えられている。例えば、哺乳類の最も古い化石種の一つとされるモルガヌコドン(図5)では脳内の嗅球、旧皮質、新皮質、小脳の拡大が見られる。

(図5) モルガヌコドン
今回、102種の哺乳類と哺乳類以外の32種の脊椎動物(は虫類、鳥類)で頭蓋骨の形成過程を比較したところ、脳を覆う5つの骨片(前頭骨、頭頂骨、底後頭骨、外後頭骨、上後頭骨)(図6)が形成される相対的なタイミングが哺乳類で著しく早いことがわかった。この5つの骨はモルガヌコドンで拡大した脳の領域の多くを覆っているものでもある。
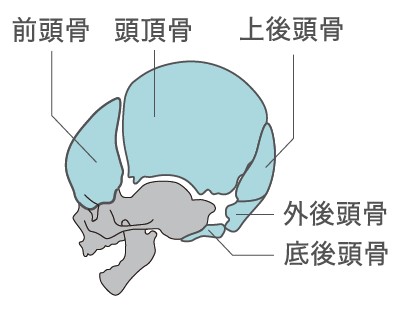
(図6) 脳を覆う5つの骨
4.大きな脳をもつ種はどこが違うのか
次に哺乳類の中での比較を行ったところ、脳を覆う5つの骨片の中でも特に上後頭骨の形成タイミングが、脳の大きさと強く関わりあっていることがわかった(図7)。哺乳類のなかでも脳の相対サイズが大きいサル類、クジラやイルカ類、モグラ類、トビネズミ類などのグループでは、上後頭骨の相対的な形成タイミングが特に早いのだ。さらに、サル類の中で最も脳の相対サイズが大きい、私たちヒトの上後頭骨の形成タイミングは、サル類の中で最も早いことも示された。哺乳類以外の脊椎動物では、上後頭骨は相対的にかなり後期に形成される。哺乳類の共通祖先でその形成タイミングが前倒しされ、さらに大脳化した一部の哺乳類でより一層早くなったと考えられる。
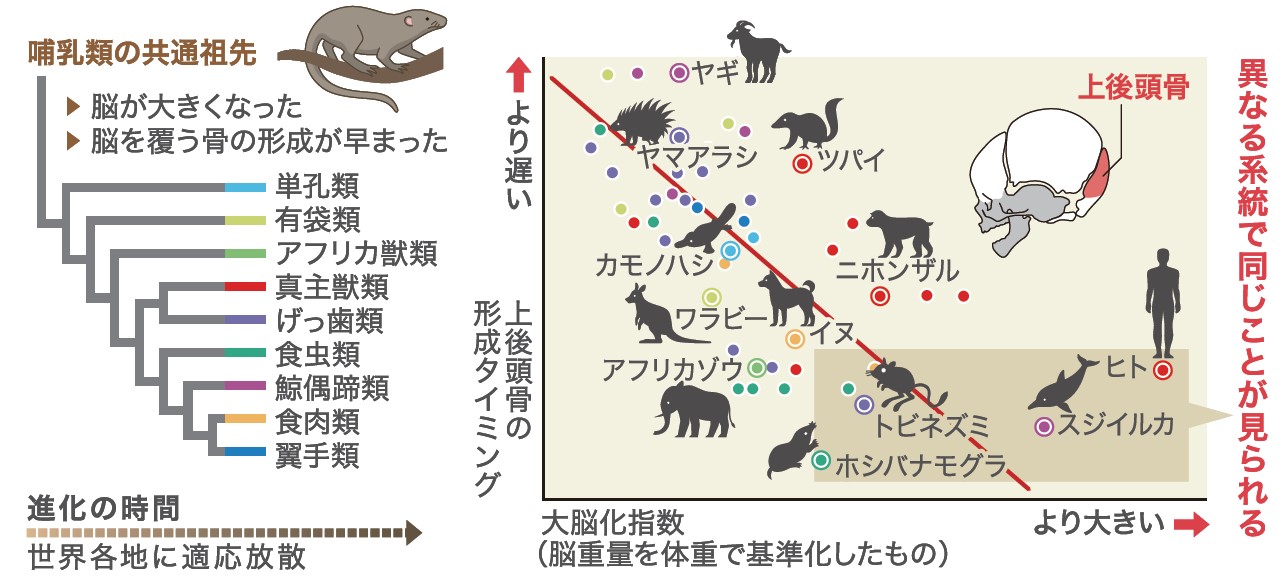
(図7) 脳の大きさと上後頭骨の形成タイミング
5.偶然と必然が重なりあって起こる形の進化
なぜ上後頭骨の形成タイミングが脳のサイズと連動しているのだろう。考えられる仮説に機能論的シナリオと多面発現シナリオの2つがある。 機能論的シナリオとは「脳が早いスピードで大きくなったのでそれを早く覆い守るため、骨の形成も早くなった」という仮説だ。直感的に受け入れやすく魅力的な仮説だが、骨形成の開始が早くなることが骨の最終的なサイズの拡大進化につながるかどうかは未検証のため、この仮説を裏づける確かな証拠は今のところない。
一方、近年の遺伝学的研究によって上後頭骨の発生と脳の発生は強く関連していることがわかっている。Lmx1bとDlx5という2つの遺伝子が胎子期のマウスの中脳と後脳の発生や軸索形成にきわめて重要な役割を果たすこともわかってきた。ヒトにおいては、これらの遺伝子の機能不全はダウン症と関わっていることが知られている。また、この2つの遺伝子が機能不全の場合、頭部の骨のうち上後頭骨の欠損や、形成の遅れが見られる。同じ遺伝子が脳と骨の形成の双方に深く関わっているのだ(図8)。このように、ある遺伝子が複数の形質の形成に影響することを「多面発現」とよぶ。脳が大きくなるという哺乳類の進化の副産物として、脳と同じ遺伝子に制御されている上後頭骨の形成タイミングも早くなったのかもしれない。
もちろんLmx1bとDlx5が哺乳類で脳と骨の双方の発生に関わるようになった背景には、機能論的シナリオがあるかもしれない。機能論的シナリオと多面発現シナリオは必ずしも二律背反の仮説ではない。ただ、現時点では少なくとも後者は脳と頭蓋骨の進化と発生を考えるうえで無視できないシナリオだろう。
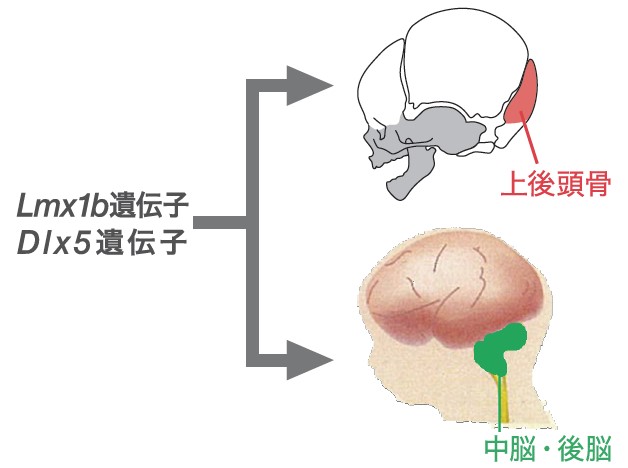
(図8) Lmx1b遺伝子とDlx5遺伝子の多面発現
両遺伝子は「上後頭骨の発生」と「中脳・後脳の発生」の双方に関わっている。
6.博物館標本の蓄積から歴史物語りを読み解く
今回の研究は、国内外の博物館で長い時間をかけて特に具体的な目的も無く収蔵蓄積されてきた希少な学術標本があったからこそ可能となった。学問の世界に限らず、「その目的は何か」とか「それは何の役に立つのか」と具体的な意義を求められることが多くなっている。しかし、博物館は「将来いつか何かの理解のために役立つはずだ」と未来に賭けながら標本を残してゆくのである。胎子の標本を残してくれた先人たちも、本研究のような活用がなされるとはよもや想像もしていなかっただろう。筆者はこれまで国内外の博物館で自分の研究活動をするだけでなく、直接自分の研究に関わらない標本の収蔵活動にも力を注いできた。それは未来の研究者に賭けているからであり、また自分の研究に使った標本を残してくれた無数の博物館関係者に恩を返したいと思ってのことである。一見さっぱり役に立ちそうにもない博物館標本を最大限に活かして標本のもつ学術価値を実証しながら、現代的な文脈でこれからも形態の進化を総合的に読み解いていきたい。そして、さまざまな視点を統合することで、私たちヒトの頭部がなぜこんなにも大きな脳をもつに至ったのか、その歴史も明らかにしたい。

小薮大輔(こやぶ・だいすけ)
2006年京都大学総合人間学部卒業。2011年東京大学大学院 理学系研究科博士課程修了。博士(理学)。京都大学総合博物館、チューリッヒ大学古生物学博物館でのポスドク研究員を経て、2013 年より東京大学総合研究博物館特任助教。














{kind=link}