RESEARCH
熱帯雨林の花ざかり
ボルネオ熱帯雨林の 一斉開花の要因を探る
日本では四季が移り変わりますが、東南アジアの熱帯雨林は一年中高温湿潤で夏の気候です。ではいつも花盛りかというと、そうではありません。通常は花が少なく、ある日突然さまざまな木々が次々に花をつける「一斉開花」があるのです。その間隔は1年以下の場合もあれば7~8年の場合もあり、周期性はみられません。季節の変化のない熱帯でどうやって木々は花の時期を知るのでしょうか。また、なぜさまざまな種の花が同じ時期に咲くのでしょうか。竹内やよいさんは、マレーシアのランビル国立公園での10年以上にわたる観察と実験から、熱帯雨林の花盛りの謎を解き明かそうとしています。
1.熱帯雨林の花の季節
日本の山では、春が来ると一斉に花が咲き始める。夏には夏の花、秋には秋の花と、毎年決まった時期に花を見ることができる。それは植物が、日長、気温などの周期性のある環境の変化に反応して、花を咲かせる時期を決めているからである。一方で、一年中温暖で多湿な熱帯の森では、花が咲く明瞭な季節がない。ある時突然、多種多様な種類の植物が次々と花をつける。この現象は「一斉開花」と呼ばれ、季節の差が明瞭でない東南アジアの低地熱帯林だけに見られる(写真1)。一斉開花といっても、すべての植物が同じタイミングで咲くのではない。ある種が花を咲かせて一週間ほど続き、ピークが過ぎたころにまた別の種が咲き始める。開花を次々にバトンタッチしていき、全体では3か月ほど続く。一斉開花はフタバガキ科をはじめとしてさまざまな系統の樹木群でおこり、その間隔は1年以下の場合もあれば7~8年の場合もあり、周期性がみられない。

(写真1) 一斉開花中のランビルの森
真っ白な林冠は開花中のフタバガキ科Dryobalanops aromatica。同じ種類の樹は疎らに生えるので、一時期には開花している木はポツポツと見られるだけである。
ボルネオにあるランビル(マレーシア、サラワク州)の熱帯雨林は、高さ50m以上にもなるフタバガキ科の樹を特徴とする東南アジアに典型的な混交フタバガキ林である。ランビル国立公園に設置されている52ヘクタールの森林調査区域には、樹木種約1200種が記録されており、これは日本全土の樹木の総種数よりも多い。同じ種の木は1ヘクタールあたり平均1個体以下であり、種類ごとの密度はとても低い。ここでは、1992年より500個体、300種以上の開花・結実フェノロジー(註1)の定期的な観察を行っており、2011年までの約20年以上に及ぶ観察で、7回の一斉開花を観測した(図1)。これらのデータから一斉開花を引き起こす要因(直接要因)と開花現象が進化してきた理由(進化要因)を探っている(図2)。
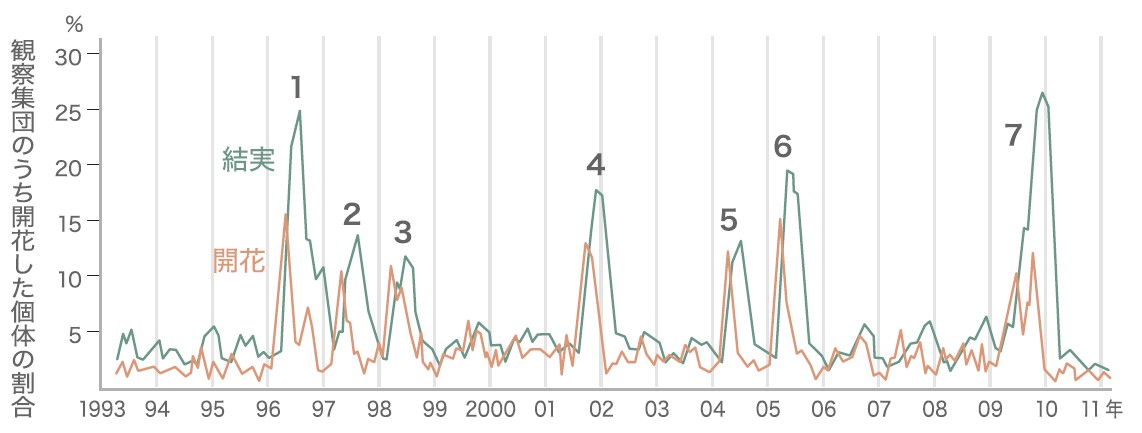
(図1) ランビル国立公園で観察された開花・結実している個体の割合(酒井ら、未発表データ)
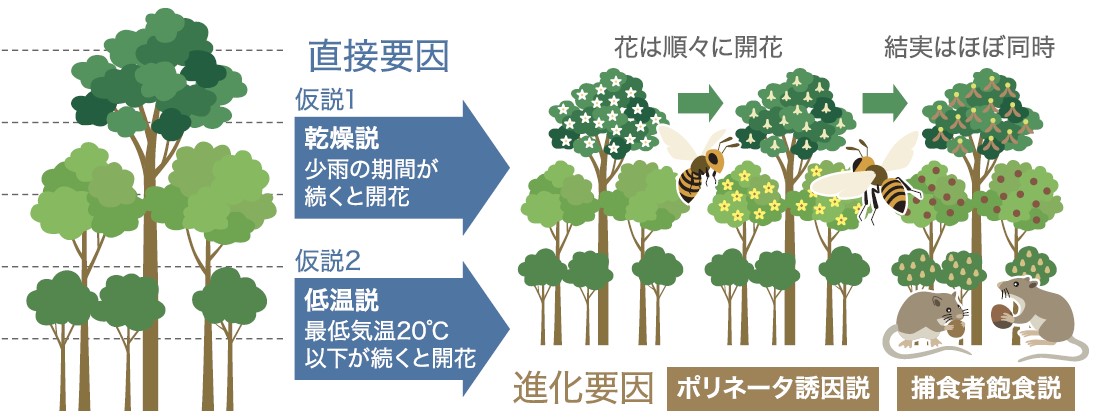
(図2) 一斉開花現象とその直接要因と進化要因
(註1) フェノロジー
動植物の示す現象と気候や気象の変化などの関係、またはその研究をする学問。生物季節(学)。
2.一斉開花の引き金
他家受粉する植物は、子孫を残すために同種の他個体と同調して花をつけなくてはならない。気象要因の変化が一年を通して乏しい東南アジア熱帯地域では、どのような気象要因が開花の引き金となっているのだろうか。二つの主要な仮説がある。東南アジア地域で不定期な間隔でおこる低温が原因とする低温説と長期(1~数か月)の少雨を原因とする乾燥説である。これまでの一斉開花の起こる時期と、最低気温及び降雨量との関係を調べた研究の結果では乾燥説を支持する例のほうが多く、低温よりも乾燥のほうが一斉開花をうまく説明できる。ランビルの観察結果でも乾燥説を支持している。しかし一斉開花は数年おきにしか起こらないのでより明確な答えを得ようと、どちらの気象要因に植物が応答しているのか遺伝子から調べることにした。
少雨による乾燥が一定期間続き、その後雨が降ると一斉に新葉を広げる「一斉展葉」現象が多くの熱帯雨林で報告されている。一斉展葉は、開花よりも頻繁にみられるが、同じように群集レベルで同調しておこる。そこで植物は乾燥をきっかけに新芽をつくり、その後の条件によって葉になったり、花になったりすると考えた。この条件が一斉開花の気象要因になるはずだ。葉か花かを決定する時期の新芽は見た目では区別できないが、発現している遺伝子を調べることで、植物がどのような状態にあるかを知ることができる。例えば、これから花になる芽であれば、花芽形成に関わる遺伝子が発現しているはずである。植物の花芽形成に関する分子生物学的な機構は、モデル植物であるシロイヌナズナで理解が進み、多くの植物で花芽形成に関わる遺伝子が共通していることが知られている。そこで、シロイヌナズナの花芽形成に関わる遺伝子の発現パタンやネットワークを参考にすることとし、この分野のエキスパートであるチューリッヒ大学の小林正樹・清水健太郎らとの共同研究により、一斉開花時の植物の反応を遺伝子レベルで明らかにする研究を行った。
3.乾燥説の遺伝子解析による検証
新芽のサンプリングは地上50メートルの林冠で行った。ランビルには高さ80メートル、アーム長75メートルの林冠クレーンが設置されており、ゴンドラに乗ってクレーンが届く範囲を3次元で移動できる(写真2)。ターゲットとしたのはフタバガキ科のShorea beccarianaという一斉開花型の種である(写真3)。2009年7、8月に雨が少ない時期が一か月以上続き、その後開花が見られた。そこで、少雨・開花の前後の芽のサンプル(図3)からRNAを抽出して、どのような遺伝子が発現しているかを網羅的に解析した。その結果、花芽ができる過程で発現が変化している遺伝子は1128個あった。そのうち、シロイヌナズナの花芽形成に関わる遺伝子の一つであるFT(FLOWERING LOCUS T :花芽形成を誘導するシグナルとなる植物ホルモン、フロリゲンをコードする遺伝子)に相当するフタバガキの遺伝子であるSbFTの発現量が、乾燥後に上昇していた(図4、図5)。一方で、花芽形成の抑制に働く遺伝子SVP(SHORT VEGITATIVE PHASE)にあたるSbSVP遺伝子の発現量は減少していた。これらの遺伝子がシロイヌナズナと同様にそれぞれ花芽形成と花芽抑制に働くことは、シロイヌナズナ遺伝子を導入し、過剰発現させることで確認している。この二つの遺伝子の変化は、見て花芽とわかる前に起こっていた。また1128の遺伝子をシロイヌナズナの持つ遺伝子と比較したところ、長期の乾燥にさらされると発現が変化する遺伝子の多くが含まれていていた。このことから、この雨の少ない期間に植物は乾燥に応答し、花芽をつくる準備をしていると考えられる。これは一ヶ月を超える少雨による乾燥が開花のきっかけとなる気象要因であることを示しており、乾燥説を裏付ける結果となった。
こうして一見まだ固い新芽であっても、植物は外界から乾燥刺激を受け取っていることや花を咲かせる状態にあることが、遺伝子の発現から明らかになった。熱帯に咲く花がどのような環境の変化を感じて開花するのかという疑問を大規模な林冠観測システムとモデル生物で得られた遺伝子の知識や花芽形成の分子生物学の手法を組み合わせ、解き明かすことができた。

上:(写真2) ランビル国立公園に設置された林冠観察用のクレーン
ゴンドラに人が乗り込み、林冠にアクセスする。林冠で葉や花に触れることができる。
下:(写真3) フタバガキ科 Shorea beccariana
いい匂いのする黄色の小さな花をつける。最大樹高は50mを超え、林冠をも超えた突出木ともなる。
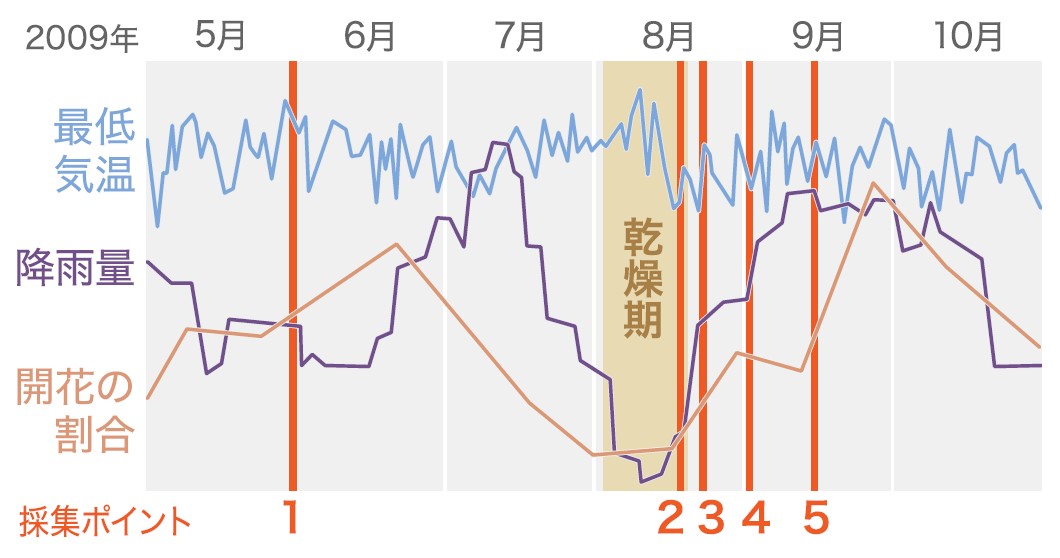
(図3)気温、降水量の関係と新芽サンプルの採集ポイント
1と2の間に乾燥がおきた。
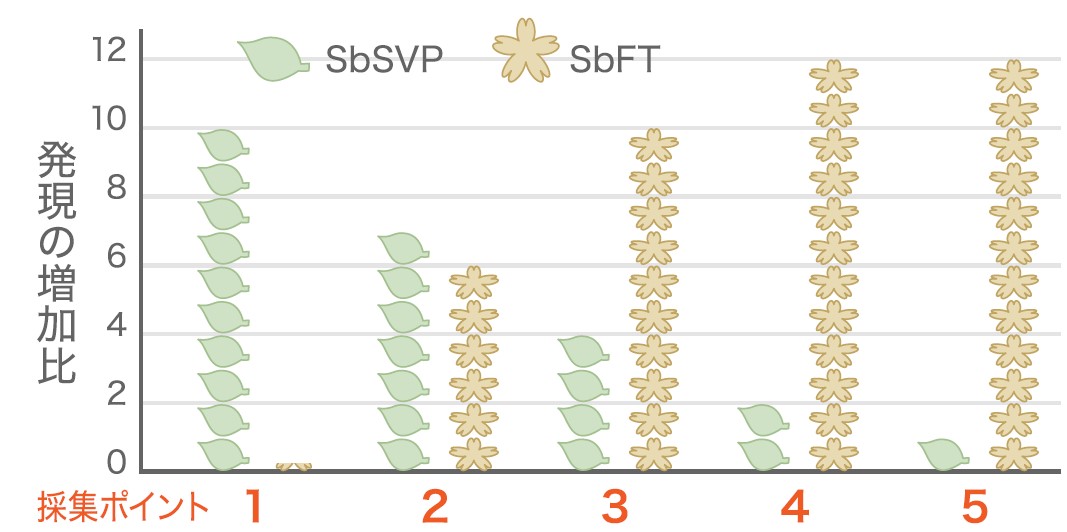
(図4) SbSVP遺伝子(花芽抑制遺伝子) SbFT遺伝子(花芽形成遺伝子)の発現量の比

(図5) シロイヌナズナへの遺伝子導入実験
フタバガキ科Shorea beccarianaで開花前後に変化が見られたSbFT(右)、SbSVP(左)の遺伝子を導入し、過剰発現させた。中央が野生型(コントロール)。右の個体は、野生型よりも早く開花した(花芽分化の促進)。一方で、左の個体は野生型よりも遅く開花した(花芽分化の抑制)(撮影:小林正樹)。
4.一斉開花の進化的な要因
一斉開花とその後の一斉結実は、花や種子を利用する昆虫、動物たちにも恩恵を与える。植物とそれを利用する動物との関係は、一斉開花を進化させることに大きく影響したであろうと考えられている。ここでは、一斉開花の進化的要因として二つの仮説を紹介する(図2)。
1. ポリネータ(註2)誘引説
一斉開花する植物には風媒花は少なく、昆虫、鳥、哺乳類などの動物媒花が多数を占める。熱帯では種あたりの数が少なく、一種で咲いても森全体ではめだたない。そこで多くの種が同調して咲けば、全体として花の数も種類も豊富になり、花粉を媒介するポリネータを誘引できるだろう。一斉開花する植物には、普段は熱帯低地林にはいないオオミツバチ(写真4)をポリネータとする種も少なくない。一斉に開花することはオオミツバチを引き寄せる威力があるのだろう。一方で、ポリネータを共有する樹種が同じ時期に咲いてしまうと、ポリネータの争奪競争になる。それを避けるために、開花期のずれがあるのではないだろうか

(写真4) フタバガキ科 Dryobalanops aromaticaの主要なポリネータであるオオミツバチ(Apis dorsata)
オオミツバチは一斉開花するとどこからともなくあらわれ、営巣する。一斉開花中は、巣の数もどんどん増えていく。そして開花が終わるとまたどこかに去っていく。
2. 捕食者飽食仮説
種子や果実を利用する昆虫や動物にとって一斉開花・結実は待望のイベントである。しかし、植物側からみれば、折角つけた種子を捕食者にすべて食べられてしまっては子孫を残すことができない。そこで普段は種子をつけず捕食者を飢餓の状態にしておき、捕食者の個体数を制限しておく。この状態で大量に結実すると、捕食者数の増加が追い付かず捕食者は飽食して食べ残すので、生き残る種子の割合が増える。この場合、結実量が多ければ多いほど食べ残しの量が増え、群集として捕食を免れる効果が高くなるだろう。
(註2)ポリネータ
植物の花粉を運んで受粉を助ける動物。主に昆虫や鳥などである。
5.捕食者飽食説を確かめる
ランビルでの20年以上にわたる開花・結実のフェノロジー観察の結果、一斉開花は毎回規模が異なることが分かってきた(図1)。例えば、2004年、2005年、2009年の群集レベルでの開花規模は、それぞれ小、中、大であった。先述の捕食者飽食仮説が正しければ、種子の生存率は開花規模の大きい2009年で高いと予測される。そこでフタバガキ科のShorea laxaを対象として種子を採集し、健全な種子と捕食され損傷している種子の割合を調べた。結果としてこの3回では差がなく、捕食者は結実の規模に応じて速やかに個体数を増やすため、規模の大きさによるメリットはみられないことがわかった(図6)。また、成木の密度と種子数の関係も解析した。木の生えている密度が高いほど捕食される種子が多い場合を「負の密度効果」と呼び、種ごとの密度が低く保たれ熱帯林で多種が共存し多様性が増す理由が説明できる。解析の結果では、2005年は密度が低いところで生存率が高く「負の密度効果」を示したのに対し、2004年は成木の密度がより高い場所で生存率が高く、2009年はどちらの傾向も見られなかった。この3回の開花からは、負の密度依存効果は熱帯林では普遍的なものではなく、一斉開花の規模によって変化していることが分かった。時期や場所などのさまざまな変動が、種多様性の維持に関わるのだろう。東南アジア熱帯林の一斉開花は、植物だけではなくそれに関わる動物を含めた種の多様性、種の構成や生態系の構造を決定する重要な役割を担っていると思われる。
今後も開花・結実の観測を続けて、熱帯林の謎に一歩でも迫りたいと考えている。異なる分野の手法や知識を積極的に取り入れることによって、ブレイクスルーにつながるような研究を目指したい。
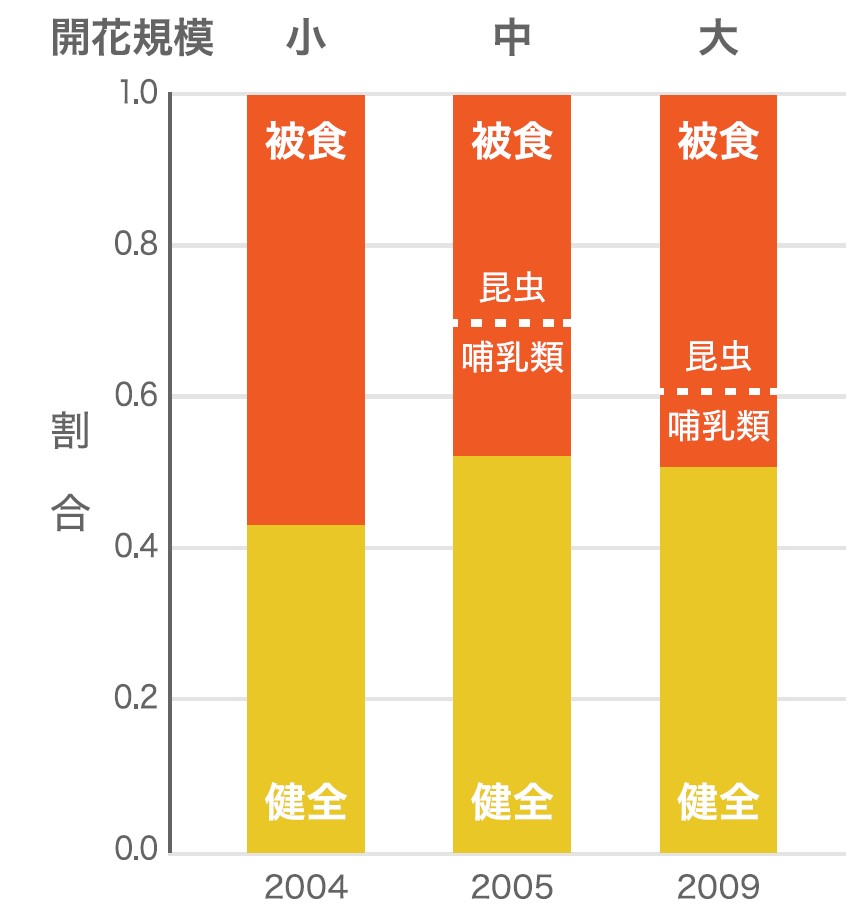
(図6) 3回の開花イベントでのShorea laxaの健全種子と被食種子(直径1cm以上)の割合

竹内やよい(たけうち・やよい)
2006年京都大学理学研究科生物科学専攻博士課程修了。総合地球環境学研究所・プロジェクト研究員、日本学術振興会 特別研究員、総合研究大学院大学・特別研究員を経て2013年より国立環境研究所生物生態系環境研究センター研究員。