RESEARCH
細胞の試行錯誤を見る
ロバスト性を支える機構を解明する
バクテリアから人間まで、生きものは遺伝子を一つ失っても、見かけ上は何も変化が現れないことが多く、これを頑強性(ロバスト性)と呼びます。例えば、大腸菌が生きていく上で欠かせないエネルギー代謝でさえ、糖からエネルギーを取り出す数多くの酵素遺伝子のどれか一つが欠けても、致命的でないことが多いのです。このとき大腸菌はどのようにして生きのびているのでしょうか。奈良先端科学技術大学院大学の森浩禎さんは、一つの遺伝子が欠けた細菌を出発点とし、そこにあらゆる組合せで二つ目の遺伝子欠失を入れ、ロバスト性が生じるしくみを解き明かそうとしています。
1.細胞のしぶとさ
生体を構成する細胞が生きていくにはアミノ酸や核酸などの成分とエネルギーの調達が欠かせない。そのために必要な酵素反応のうち連なって一つの役割をする場合を代謝経路と呼ぶ。その中で最も中心的な働きをするのが、解糖系、ペントースリン酸経路、TCA回路である(図1)。これはもちろん、バクテリアからヒトまですべての生きものに存在するが、奇妙なことにこの経路に存在する酵素遺伝子のうちいずれかを失っても死なないという現象がしばしば見られる。この性質を頑強性(ロバスト性)という。生きものはなかなかしぶといのである。重要な代謝経路が遮断されているはずの細胞がなぜ生き延びるのだろうか。
我々は大腸菌を用いてこの問いの答えを探すことにした。人の腸内に共生する通常無害な細菌であり、分子遺伝学のモデル生物とされ、遺伝子の構造や機能の解明のほとんどはこの細菌で行われたと言ってもよい。20世紀末に生命現象を知るには生物の持つDNAのすべて、つまりゲノムを知る必要があると考えられるようになった時も、いち早くその配列解析が始まり、1997年にアメリカと日本のグループにより解読された。約4100のタンパク質を作り出す遺伝子から構成されていることが明らかになった。こうして「全体を見る」ことができるようになり、細胞がどのように生きているかを知るための研究ができるようになったのである。大腸菌は一つの細胞が個体であること、遺伝子の数も多くないので細胞の機能を丸ごと見るのにうってつけの対象である。
中心代謝系の遺伝子群を含めて、すべての遺伝子がいつどのくらい発現するかを測定する技術はある。さらに、代謝反応の結果、細胞の中で合成されたり、消費されたりする物質の量の測定もできる。そこで、これらの測定値を基にコンピューターで生命現象を再現し、実際の細胞との比較を通して生きもののしくみを明らかにするシステム生物学という分野が生まれている。そこで私たちは重要な遺伝子を失った細胞の中で何がおきているのか、どのような補償が働いてロバスト性が生じるのかをシステム生物学で解析しようと考えたのである。それには、まずそれを調べる実験系をつくる必要がある。ある遺伝子を欠いても死なないという現象を知るためには、更にもう一つの遺伝子を壊し、二つの遺伝子がはたらかないという株を全遺伝子について作ればよい。二重欠失株ライブラリーづくりである。
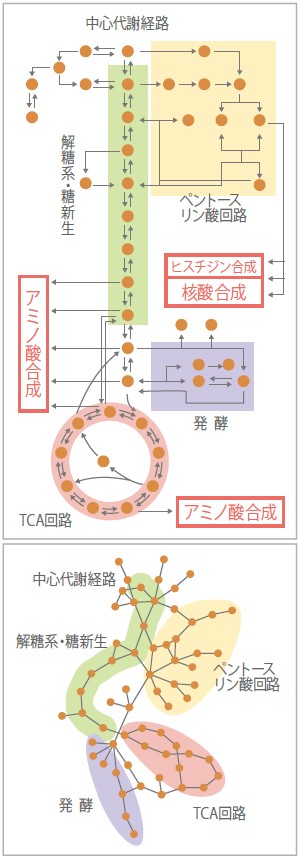
(図1)中心代謝経路
解糖系、TCA回路、ペントースリン酸系、発酵などの生存に重要な代謝経路からなる。上は生化学の研究から調べられた教科書にもよく登場する経路図。下は上の図と同じであるが、既知の代謝反応産物を結合させただけの経路図。人の見やすさを省くことで見えてくることもある。
2.二重欠失株ライブラリーの作製
実は私たちのグループでは全遺伝子クローンライブラリーと遺伝子欠失ライブラリーを構築し、それを用いてシステム生物学を考えてきた。ところが、欠失株ライブラリーを作製してみると多くの遺伝子が欠失しても生存に関わらないということが分かってきたのである。その背景には様々なしくみが考えられる。例えば、予め同じ酵素遺伝子(アイソザイム)をいくつか用意し、経路を多重化し一つが使えなくなったときバイパスすることなどである。
本来の経路を逆流させて必要な物質を作り出すなど回路が破損してから新しく経路自体を設計しなおすこともあるだろう(図2)。生命体はこれらさまざまな可能性を組み合わせ、突然変異やウイルスなどの感染による遺伝子の破壊、生育環境の急激な変化など厳しい中を生き延びてきたに違いない。
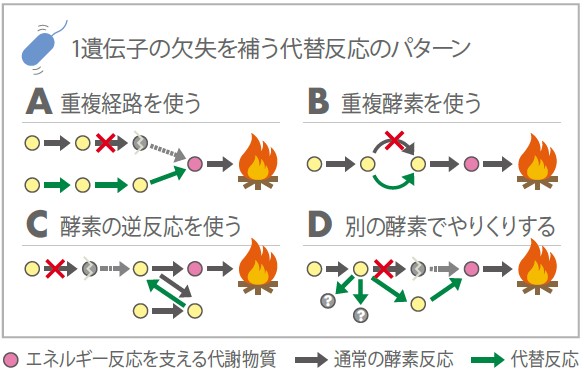
(図2)遺伝子欠失がロバスト性を示す例
そこで、前述のように、二つの遺伝子を同時に破壊して生育を調べることにした。一つの経路を遮断したときに細胞が取るであろうもう一つの経路を遮断すれば死んだり、あるいは生育が悪くなったりするだろうと考えたのだ。
大腸菌の約4000遺伝子すべてについて二重欠失をさせると、4000 × 4000で1600万通りの欠失が必要となり、これをすべて作るのは不可能と思われるかもしれない。しかし、大腸菌には雌雄があり、雄株にはF因子と呼ばれる小さなDNAがある。接合によって雄株から雌株へとF因子を用いて遺伝子を移すことできる(この性質があるので、分子遺伝学のモデル生物になったとも言える)。そこで、ある遺伝子欠失を持つ雄株と別の欠失を持つ雌株を混ぜるだけで、たやすく二重欠失株を作ることができるのだ。私たちは3年かけて、4000個の遺伝子を一つずつ欠失させた菌株コレクションを雄雌二種類作った(図3A)。そして、手の平サイズの寒天培地の上での一回の操作で、雄株1種 × 雌株1500種の組合せで二重欠失株を作ることができるようにした(図3B)。二年あれば大腸菌遺伝子のすべてについて二重欠失株を立ち上げることができる状態になったのである。全遺伝子の組合せによる解析実験が可能になったことは大きな前進である。
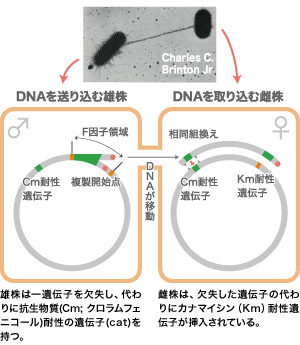
(図3A)二重欠失株の作成
雄株では、F因子からゲノムが複製され、雌株に送り込まれる。
雌株では雄株の欠失遺伝子部分との相同組換えが起こり二つの遺伝子がない代わりに、Cm、Kmの二種類の抗生物質を加えた培地で生育できる二重欠失株になる。

(図3B) 二重欠失株の検出
二重欠失株の大腸菌の生育を調べる装置。手のひら大のプレート(右)に1500株の大腸菌を育てて、コロニーの大きさを市販のスキャナでスキャンして、コンピュータ処理すると1日に最大74,000 コロニー処理できる。
3.二重欠失株から見えるロバスト性
遺伝子欠失株の研究を通じて、遺伝子産物が作り出す細胞内の代謝経路は非常にダイナミックに組換えられ、その時々の状況に応じて最適化されていると考えるようになった。教科書では、解糖系はグルコースからピルビン酸までを代謝する一本のまっすぐな経路として描かれ、その下に環状に描かれたTCA回路からはアミノ酸などの合成へと枝が広がっている。しかし、細胞内の酵素反応をブロックとして組み合わせると別の見方ができる。代謝物質を丸印で描き、酵素反応を線で結ぶと、多くのループ経路が存在するのだ(図1)。このようなループがあれば、一つの酵素反応を失っても、他の道を辿って欠けた反応の生成物を供給できる。アイソザイムによって、二重化されている経路もある。代替経路は正常のときはあまり使われていないので、一段階ずつ酵素反応を調べる研究では見つかりにくい。今回のように代謝全体を見渡して初めてその存在が見えてくるのである。「全体を見る」研究の重要性はここにある。常に使われているのだが、反応の頻度が少なく主となる経路が使えなくなると処理量が増える場合もあるだろうし、細胞を維持する経路は状況に応じて使用頻度が変わる「ゆらぎ」や「遊び」を含んでいるのだ。それがロバスト性を支えているのだろう。
実際に二重欠失株の実験から見つかった代替経路がある。大腸菌には、呼吸によるエネルギー生成などの酵素の活性中心として働く鉄硫黄クラスター(Fe−Sクラスター)の合成経路が二つ知られている。トロント大学との共同研究で、この二つの経路Suf経路とIsc経路が補完的であることを証明した。二重欠失株でどちらか一方の経路で二つの遺伝子を欠失しても生育できるが、それぞれの経路から一つずつ欠失させると致死になったのである(図4)。Suf経路とIsc経路はFe-Sクラスター形成における多重化された経路で、片方が使えなくなっても、もう片方が残っていれば事足りるのだ。この研究では、従来Suf経路の遺伝子として知られていたもの以外に、Isc経路の遺伝子と同時に欠失させると致死になる新規の遺伝子が見つかった。二重欠失株による解析ならではの結果である。
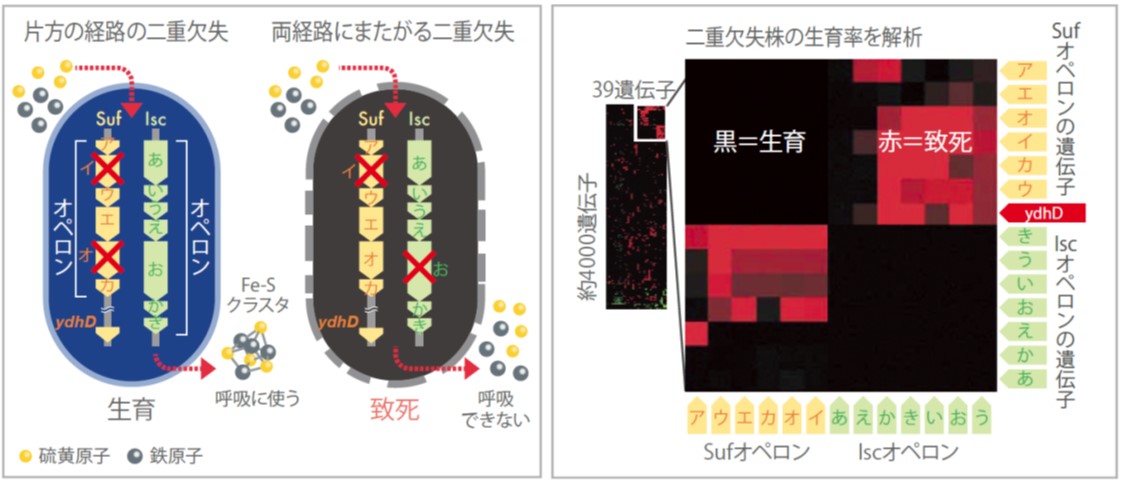
(図4)Fe-Sクラスタ合成に関わるSuf経路とIsc経路の遺伝子の二重欠失の関係
Suf経路同士、Isc経路同士の遺伝子を二つ欠失しても生育できるが、それぞれ一つずつを欠失し両方の経路が遮断されると致死になる。
慶応大学との共同研究では、中心代謝経路に未知の反応経路を発見した。トランスアルドラーゼはペントースリン酸回路でセドヘプツロース7−リン酸(S7P)という糖を分解するので、この酵素の二つのアイソザイム欠失株はS7Pを分解できず、菌体にS7Pが蓄積するはずである。ところが代謝産物の量を測定すると蓄積はしておらず別の代謝物ができていた(図5)。この分解には、正常時にはもっぱら解糖系で働く二つの酵素が関与していることがわかった。S7Pが蓄積したことが引き金となってはたらくようになったのだろう。つまり状態が変化すると既存の酵素によるバイパスができるのだ。中心代謝経路は古くから知られており、研究され尽くした経路と考えられてきたが、まだ面白いことが隠されているのである。二重欠失株とその表現型解析にメタボローム解析(代謝産物の網羅的解析)やトランスクリプトーム解析(遺伝子発現の網羅的解析)を組み合わせると、新規の経路の発見が可能なのだ。このような個別の解析によって様々な代替経路のパターンを蓄積し、方法を確立していくことが今後の解析の弾みとなるはずである。
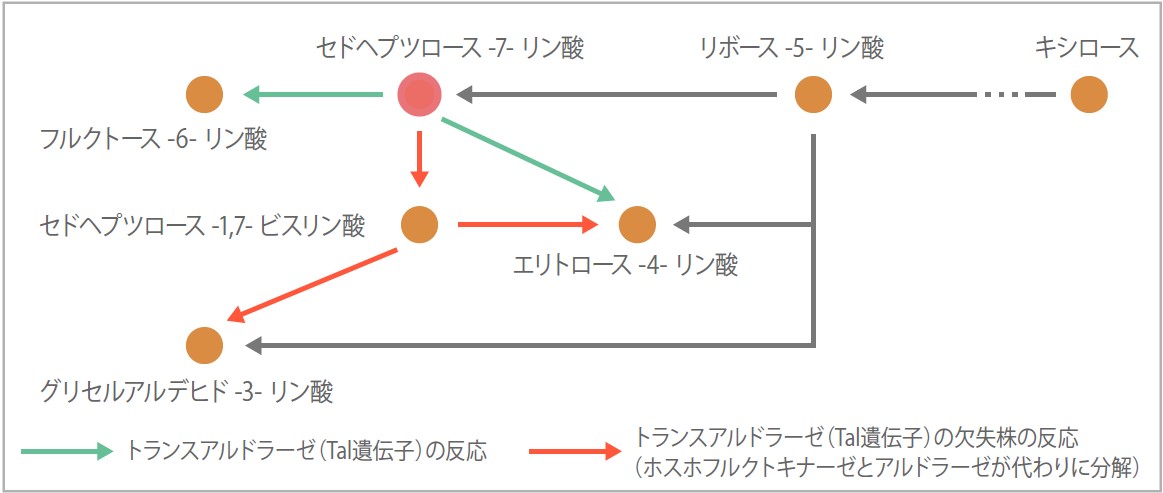
(図5)トランスアルドラーゼ欠失株で発見されたバイパス
正常の株ではセドヘプツロース-7-リン酸は、緑の経路で分解されるが、トランスアルドラーゼがないと解糖系の酵素(ホスホフルクトキナーゼ、アルドラーゼ)が代わりに分解し、赤の経路で反応がおこる。
4.生命とロバスト性
大腸菌というたった一つの細胞からなる生物が環境の変化、遺伝子の損傷などに対して抵抗し、適応するロバスト性を実感した。ロバスト性は、固く硬直した強さではなく、いくつかの可能な経路の存在つまり遊びを許し、その中をゆらぎながら環境や状態に最適な系を選んで使うしなやかで柔軟な強さである。生命体が38億年にもわたって続いてきた理由は、生命体の最も基本的な性質としてロバスト性があるからだろう。技術的には可能になったとはいえ、1600万の組み合わせの意味を知り尽くすのはまだ遠い先のことだ。しかし、生きている細胞の理解が始まっているのである。それがわかって初めて細胞を巧みに用いる医学や産業への応用ができるのである。細胞の全体像を描くことを夢見て、冷凍庫いっぱいの菌株と計算機に蓄積するデータとの格闘を続けている。
森 浩禎(もり・ひろただ)
1985年京都大学理学研究科修了後、熊本大学医学部助手、 京都大学ウイルス研究所助手を経て、1996年より奈良先端科学技術大学院大学バイオサイエンス研究科教授。理学博士。














