RESEARCH
わが子の食草を探す準備はさなぎの中で
味覚受容体遺伝子がむすぶ
化合物と産卵行動
ナミアゲハの幼虫はミカン科の葉しか食べないので、親はそこに確実に産卵しなければなりません。食草には産卵刺激物質が含まれ、雌チョウの前脚ふ節の感覚子にそれを識別する「味覚受容体」があるのです。この「味覚受容体」は、いつどこで作られ、チョウの産卵行動にどのような影響を与えているのでしょうか。
1.アゲハチョウと食草
地球上で最も多様化した生物は昆虫であり、その過半数が植食性である。昆虫の中で比較的新しいグループとされる鱗翅目の幼虫は大部分が植食性で、多くは特定の植物の葉しか食べない。アゲハチョウの仲間も、狭い範囲の植物しか食べず、しかもふ化直後の幼虫は体が小さくて移動能力が低いので、メス成虫が正確に植物種を識別して産卵しなければ、次世代は育たない。まずこの仕組みを知りたい。しかもその仕組みが変化して、食性が変わると、そこから種分化が起きるのではないかという問いをたて研究を始めた。
アゲハチョウのメス成虫の産卵行動は、飛ぶことから始まる。主に視覚情報を頼りに葉にとまり、葉を前脚でたたく。これを「ドラミング」と呼ぶ。前脚ふ節には化学感覚子があって、葉の表面に存在する化合物を感じ取り、その組み合わせによって植物種を識別し、適切と判断すると卵を産む。その後その場を飛び去り、一連の作業を繰り返すのである。ナミアゲハについては、主な食草であるミカン科植物から、化合物10種が「産卵刺激物質」として同定されており、これらを混合すると生葉と同程度に産卵行動を誘発する(図1)。

(図1) ナミアゲハの産卵実験
ミカン葉から抽出した産卵刺激物質を塗布した人工葉に産卵している。
一方、化合物を識別する受容体の研究は、私たちの研究開始時にはまったく行われていなかった。ショウジョウバエの味覚感覚子では、感覚子内部へ数個の味覚ニューロンが軸索を伸ばしていること、その細胞膜中に7回膜貫通型タンパク質(7TMP: 7 transmembrane protein)である味覚受容体が存在することが知られており、化合物を認識しているとされている。チョウの前脚ふ節にある感覚子は、ショウジョウバエの味覚感覚子と同様の構造を持っているので、アゲハチョウのふ節感覚子でも、7TMP味覚受容体が働いていると予想できる(図2)。これが食草の認識と産卵行動に重要な役割を持っているだろうと考え、ナミアゲハの味覚受容体遺伝子の探索と機能解析に取り組んだ。
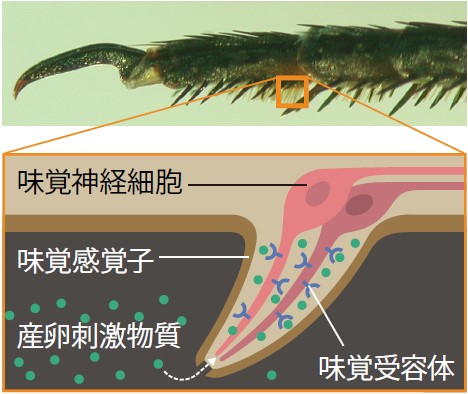
(図2) ナミアゲハの味覚感覚子
ナミアゲハの味覚感覚子は、メス前脚の先端のふ節の毛のように並んでいる。味覚受容体は、味覚感覚子の中の味覚神経細胞の表面にある。
2.味覚受容体遺伝子を追いかけて
味覚受容体遺伝子は、発現量が極端に少ないため、他の生物でもタンパク質の機能が解明されているものはまだ少ない。しかも昆虫の化学受容体遺伝子は、遺伝子間の塩基・アミノ酸配列の類似性が極端に低いので、他の生物の受容体遺伝子を手がかりにはできない難点がある。そこで、メス成虫ふ節で発現する遺伝子群を片っ端から調べることにした。メス成虫ふ節のcDNAライブラリーをつくり、約1万個の配列を決定してアミノ酸配列を推定した(註1)。その中で7回膜貫通領域を含む配列を持つものをコンピュータ上で探し、候補遺伝子を絞り込んだ。そしてついに味覚受容体候補の7TMP遺伝子を1つ見つけることができた。得られた7TMPは、メス成虫のふ節で発現していることはわかった(図3)が、7TMPには神経伝達など化学受容以外の働きを持つものもたくさんあるので、本当に食草の認識に関わる味覚受容体かどうかを確認しなければならなかった。そこで「カルシウムイメージング法」を用いた。まず味覚細胞ではない培養昆虫細胞でナミアゲハ味覚受容体候補7TMPと発光クラゲから見つかったイクオリン(aequorin)という発光タンパク質を発現させる。その細胞を化合物で刺激したとき7TMPが応答すると発光タンパク質がカルシウムに反応して光るという仕組みである。私たちが得た7TMPは、産卵刺激物質の中のシネフリンを与えた時だけ発光した(図4)。一か八かの実験だったが、2年間大量の遺伝子を調べる実験を許されたおかげで、産卵刺激物質シネフリンを認識するナミアゲハの味覚受容体遺伝子を手にすることができた。

(図3)味覚受容体遺伝子の発現確認の実験
発見した味覚受容体遺伝子が、チョウの体のどこで働いているか遺伝子を増幅して調べた。電気泳動のシグナルは、メス前脚ふ節にだけ検出された(左1列目のバンド)。比較に用いたリボソーム蛋白質遺伝子は全部の器官にみられるので、味覚受容体遺伝子がメスの前脚ふ節でだけ働いていることが確認できた。
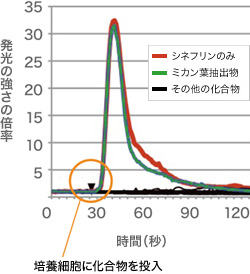
(図4)「カルシウムイメージング法」で得られた発光の比較
味覚受容体遺伝子を組み込んだ細胞は、シネフリンだけで刺激した場合でもミカン葉抽出物でも同じ強さで発光する。つまり、この味覚受容体は、ミカン葉に含まれるシネフリンだけに反応していることがわかる。(縦軸は、発光の強さの相対的な倍率)
3.チョウのRNAiに挑戦する
遺伝子のはたらきを調べるには、その遺伝子のメッセンジャーRNAの発現を阻害する二本鎖RNA(dsRNA)を生体に導入するRNA干渉(RNAi)がよく使われる。鱗翅目昆虫では、dsRNAの注射によって遺伝子の発現量を抑えることはできるが、その効果が持続せず、この方法は難しいとされてきた。しかし私たちは、ナミアゲハ7TMPの発現量がピークになる前にdsRNAを導入し、発現レベルが低い状態にすれば、阻害効果がでるのではないかと考えた。そこで成虫だけでなく蛹からもふ節を取り出し、日を追って7TMPの発現量を調べてみると、羽化前日の蛹で発現量がピークとなることがわかった。そこで、羽化日を0日として-1, -3, -5, -7, -9日の蛹にdsRNAを注射で導入し、羽化直後の発現量を調べてみた結果、蛹化後の早い時期にdsRNAを注射するほどこの遺伝子の発現を効果的に抑制できる事がわかった。しかし、蛹化後早い時期は体が柔らかく、注射によるダメージが大きいので、蛹の死亡率がやや高まる。十分な遺伝子発現抑制効果は得られ、蛹の死亡率は低い時期として、羽化5日前が最良と考え、以後のRNAi実験では、羽化5日前の蛹にdsRNAを注射した(図5)。

(図5) 蛹へのdsRNA溶液の注射
羽化前5日のメスの蛹に対し、胸部と腹部の境界部分に、斜め後方から細い注射針を突き刺す。この頃蛹の内部では、このあたりに脚ができはじめている。
次に、RNAi法を用いて7TMP遺伝子の発現を阻害したチョウの前脚を用い、電気生理学的解析を行った。感覚子にある神経細胞(ニューロン)が認識する化合物で感覚子を刺激すると、中枢系へ送られる電圧が変化(神経発火)するという事実を利用した。シネフリン受容体であるナミアゲハ7TMPの発現をRNAiで抑制すれば、シネフリンによる刺激で観察される神経発火が減少するはずである。実際にRNAi処理を施したアゲハチョウ(RNAiチョウ)の前脚では、シネフリンによる刺激に対してだけ神経細胞の応答が有意に減少し、スタキドリンやカイロイノシトールといった別の化合物での刺激に対しては、応答は減少しなかった(図6)。
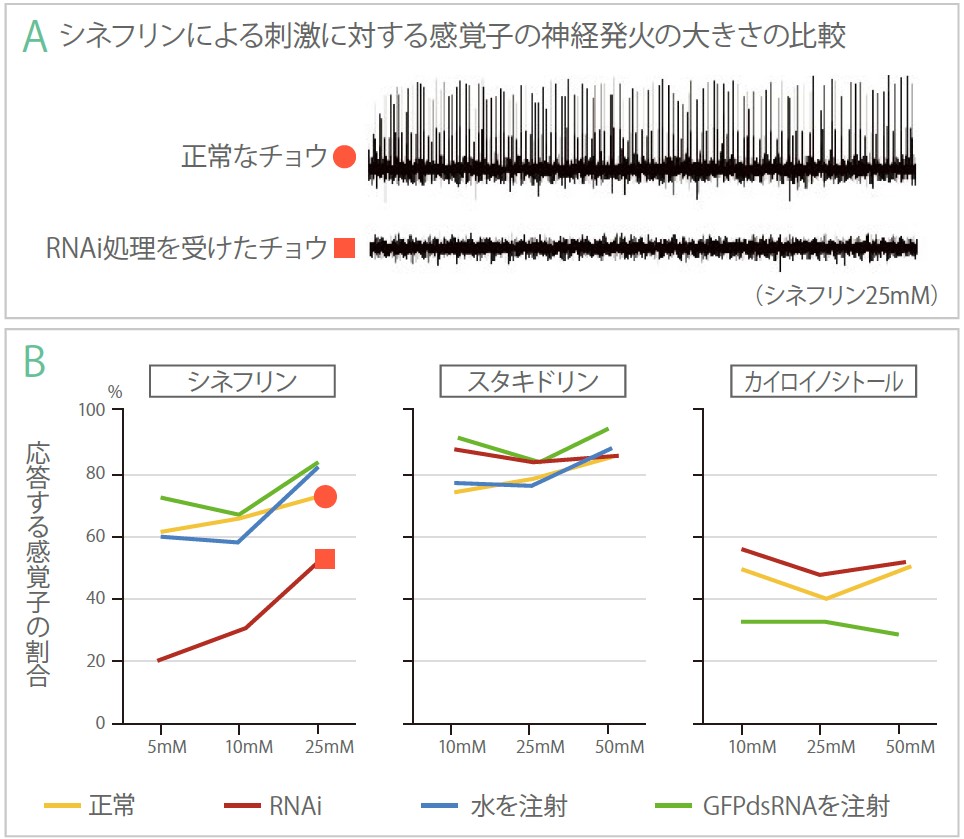
(図6) RNAi処理を受けたチョウによる電気生理の実験の結果
A. 正常なメスの前脚ふ節の味覚感覚子(上)は、シネフリンに対して神経発火を示す。一方、RNAi処理を受けたチョウの感覚子(下)は、応答が消えている。
B. A.で示した神経発火を産卵刺激物質を変えたり、濃度を変えたりして応答する感覚子の割合を調べた。RNAiによる応答の減少はシネフリンの時にだけ見られる。
4.味覚受容体がチョウの産卵行動を変化させる
次に、RNAiチョウを用いて自由飛翔による産卵実験を行った。チョウが飛び回るのに十分な広さがある約50センチ四方の容器の中にアゲハチョウを放し、プラスチック製の人工葉を提示、チョウが自ら人工葉に接近・ドラミングし、産卵するか否かを観察するという気の長い実験である。ミカンの葉から抽出した化合物や、10種類ある産卵刺激物質のうち水に溶けやすい5種類を混ぜて人工葉に塗布した場合、生の葉を触らせた場合と同等の高い産卵活性を示す。水溶性の産卵刺激物質のうち、シネフリンとカイロイノシトールの2種類だけを混ぜた場合にも、生の葉の約70%の産卵活性が見られる。しかし、どの産卵刺激物質も、単独では全く産卵活性がない。そこで、RNAiによってシネフリン受容体であるナミアゲハ7TMPの発現を抑制したチョウは、シネフリンを認識できないのだから、シネフリンとカイロイノシトールの混合溶液に触らせても、カイロイノシトールのみでの産卵実験と同じように活性なしとなるはずと考えた。
実際に、容器内で自由に飛翔するRNAiチョウにシネフリンとカイロイノシトールの混合液を付着させた人工葉を提示したところ、飛翔行動とドラミング行動は、通常のチョウと全く変わらないのに産卵活性が有意に低下した。この結果をまとめた表を文末に示すので、ご参照いただきたい。通常、卵を産む場合には、ドラミングの直後に迷うことなく腹部を曲げて卵を産み付け(動画1)、産まない場合はあっさりと飛び去ってしまう。水や単独の産卵刺激物質溶液を塗布した人工葉など産卵刺激がない場合、ドラミングの直後にさっと飛び去る。チョウはドラミングによって瞬時に卵を産むか否かを判断するのである。それに対し、シネフリンとカイロイノシトールの混合溶液に触れたRNAiチョウは、普通のチョウとは比べものにならないくらい長時間ドラミングを繰り返し、さんざん悩んだあげく卵を産まずに飛び去ってしまう(動画2)。手前味噌だが、この動画は研究者の目で見ての傑作である。RNAiによる遺伝子の発現阻害では、ノックアウトと違って発現量を0にすることはできないので、RNAiチョウもほんの少しだけシネフリンの味を感じることができているのではないだろうか。しかし産卵活性を引き起こす閾値には達しない程度の刺激であるため、悩んだ末に産まないという判断をしているのではないかと思われる。この様な"迷い"は、RNAiという実験手法だからこそ観察できた行動であろう。
以上見てきたとおり、私たちが発見した受容体遺伝子は、産卵刺激物質の一つであるシネフリンを特異的に認識し、産卵行動の誘発に関与することで、アゲハチョウの食草認識において重要な役割を担っていることがわかった。
私たちは、チョウの仲間が食草を転換することを出発点として棲み分けが起こり、種分化につながったのではないか、という仮説の証明を目指している。今後、複数種のアゲハチョウから産卵刺激物質の認識に関わる遺伝子を複数見つけ、その変化を比較することで、食草を通してのアゲハチョウの進化を見て、できることならそれを進化一般の理解につなげていきたいと考えている。
(動画1)シネフリンとカイロイノシトールの2種類の化合物を浸した人工葉による産卵実験:正常なチョウ
正常なチョウは、葉にとまるとドラミングの直後に腹部を曲げて、プラスチックの葉に卵を産みつけて、飛び去る行動を繰り返す。
(動画2)シネフリンとカイロイノシトールの2種類の化合物を浸した人工葉による産卵実験:RNAi処理を受けたチョウ
RNAiを受けたチョウは、飛翔・プラスチックの葉への着地・ドラミングまでは正常なチョウと同様であるが、その後ドラミングを長時間行うが、卵を産まずにいったん飛び去る。しかし、気になるのか、戻ってきてまたしつこくドラミングするが、結局産卵しないで飛び去る様子が観察される。
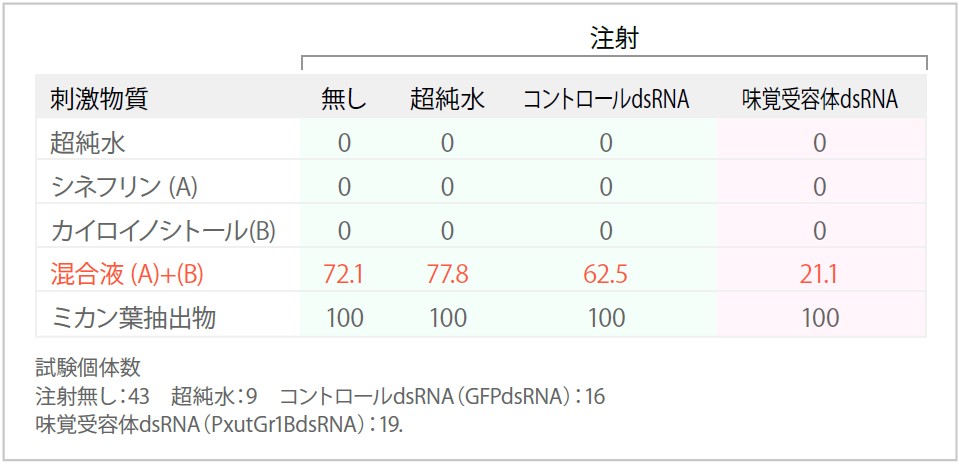
(表)味覚受容体遺伝子(PxutGr1)dsRNAの注射による産卵行動への影響(産卵率%)
尾崎克久(おざき・かつひさ)
2000年弘前大学(岩手連大)博士課程修了。博士(農学)。農林水産省果樹試験場りんご支場生物系特定産業技術研究推進機構ポスドク、JT生命誌研究館奨励研究員を経て2005年より同研究員。