RESEARCH
細胞のダイナミズムが生み出す
左右相称の形
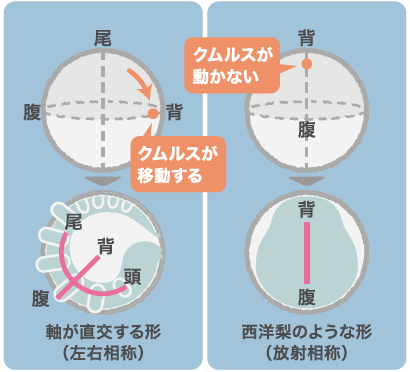
多くの動物は体に頭尾(前後)軸と背腹軸とをもっています。オオヒメグモの初期胚でパッチトと呼ばれる遺伝子をはたらかなくしたところ、背腹軸の向きを決めるクムルスが動かず西洋梨のような形になったのです。西洋梨形の胚では尾の領域が形成されなかったことから、頭尾軸の形成と背腹軸を決めるクムルスの動きには関わりがあるに違いないと考えました。
1.細胞の移動と直交する二つの軸の形成
どう見てもおかしな形の卵に気がついたのは3年も前のことである。ある特定の塩基配列をもつ二本鎖RNAを注射(註1)した母グモから生まれた卵が、西洋梨のような形になっていたのだ。どうやらクムルスが動かなかったらしい。私たちは動物の体づくりの基本となる直交する二つの軸、前後(頭尾)軸と背腹軸が、どのようにして形成されるかをオオヒメグモを用いて研究している。それまでの研究で、クムルスには、丸い卵の放射相称性を壊し、背腹の軸を形成するために重要なはたらきをする細胞集団が存在することを明らかにしていた(関連記事へ)。この細胞集団は背側を誘導する因子Dppを分泌する。正常発生では、クムルスの細胞集団は卵の北半球を覆う細胞シートである胚盤の基底面を北極から赤道領域へと移動し、移動した側に背側を誘導する。つまり背腹の軸は北極と南極を結ぶ軸と直交することになる(図1)。ところが梨形の卵では、細胞集団は移動せず、北極に留まったまま背側を誘導した。このために、背腹軸が北極と南極を結ぶ向きとなり、放射相称性は壊されなかったのだ。
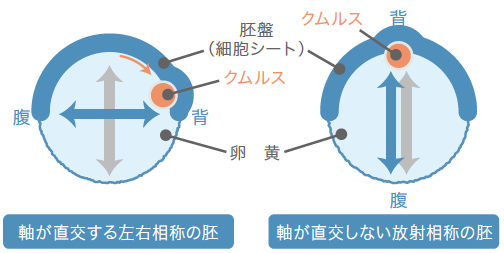
(図1)二つの軸を直交させるクムルスの移動
多くの動物は直交する前後軸と背腹軸をもつ。しかし軸を直交させる仕組みが解明されている例はほとんどない。オオヒメグモではDppを分泌するクムルスの細胞集団の移動が二つの軸を直交させる現象であると考えられたため、この仕組みを解くことを目指していた。しかし、dpp を含む数十の遺伝子の機能を抑える実験を行ったが、細胞集団の移動が異常となる遺伝子は見つからず、仕組みを解析する手がかりすら、なかなか得られなかった。そのような中で、西洋梨形の卵を見つけたことは大きな進展へとつながった。
(註1) 二本鎖RNAを注射
細胞質に存在する二本鎖RNAと相補的な塩基配列をもつmRNAが分解されるRNA干渉と呼ばれる現象を利用し、選択的に、任意の遺伝子発現を抑制する場合に用いる手法
2.ヘッジホッグ・シグナルはクモ胚の前後軸形成にはたらく
卵が西洋梨形となった実験で母グモに注射したのは、パッチト (patched:ptc )と呼ばれる遺伝子の塩基配列をもとに合成した二本鎖RNAである。注射により、生まれた卵ではptc 遺伝子の機能が抑えられる。ptc を抑える実験を行うきっかけとなったのは、クモ胚の初期発生にはたらく遺伝子を探しているうちに、ヘッジホッグ (hedgehog:hh ) と呼ばれる遺伝子の機能を抑えた卵に大きな異常を見つけたことである。Hhは細胞外に分泌されるシグナル分子であり、その受容体が細胞膜に存在するPtcである。いくつかの動物種でHhやPtcのはたらきに関して研究が進んでいる。PtcはHhと結合していないときには、スムースンド(Smoothened:Smo)と呼ばれる膜タンパク質の機能を抑制する。ところがHhが結合すると、PtcのSmoに対する抑制が解除され、SmoはHhシグナルを細胞内に伝達する。Hhシグナルは、脊椎動物の四肢や中枢神経系、節足動物の体節などの形成に重要な役割を果たすことが知られている。
オオヒメグモの正常な胚発生では、クムルスの移動後に放射相称な形の胚盤が左右相称な形の胚帯になる。hh 遺伝子の機能を抑えた胚ではクムルスの移動までは正常に行われているように見えたが、その後、胚盤は胚帯へと成長せず縮んでしまった。分かってきたのは、正常胚であれば胚盤の周縁部(卵の赤道付近)で発現するはずの遺伝子が発現せず、反対に、胚盤の中心領域(北極付近)だけで発現するはずの遺伝子が胚盤全体(北半球)で発現していたこと(図2中央)。胚盤の周縁部で発現するのは頭部形成に関わる遺伝子で、中心領域で発現するのは尾部形成に関わる遺伝子である。hh 遺伝子の機能を抑えた胚は頭部を欠き、全体が尾部化してしまったのだ。
では、受容体のPtcがはたらかないとどうなるか? という疑問から行ったのが冒頭の実験である。ptc の機能を抑えた胚を西洋梨形になる前の胚盤の時期に解析すると、hh の場合とは対照的に、中心領域で発現するはずの尾部形成に関わる遺伝子の発現が全く検出されず(図2右)、周縁部で発現する頭部形成に関わる遺伝子の発現領域が拡大していた。観察された異常がhhとptc それぞれをはたらかなくした胚で対照的だという結果は、Hhシグナルの伝達におけるPtcのはたらきを考えると矛盾なく説明できる。それはPtcがはたらかないと、Smoが常に抑制を解除された状態となり、たとえHhが無くてもHhシグナルが細胞内に伝わり続けると考えられるからだ。こうして、Hhシグナルがクモ胚の前後軸形成にはたらく遺伝子ネットワークの中のおおもとに位置することが分かってきた。多くの発生現象で重要な役割を果たすHhシグナルだが、こんなにも初期の胚における役割が明らかになったのは初めてであった。
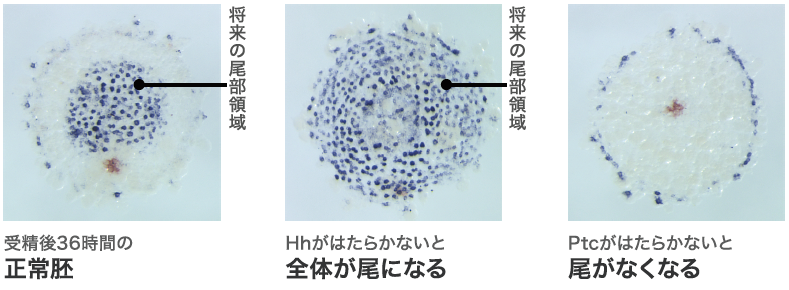
(図2) Hhをはたらかなくした胚とPtcをはたらかなくした胚
3.前後軸形成と背腹軸形成は協調している
このように、私たちはHhシグナルがクモ胚の前後のパターンを形成するという新たな事実を発見した。しかし、ptc の機能を抑えた胚で、なぜクムルスの細胞集団の移動が起こらないのか? という問いへの答えはまだ出ていない。そこであらためて正常胚で、細胞集団の移動と胚盤における遺伝子発現の変化を観察したところ、尾部の形成を示すDelta 遺伝子の発現が胚盤の中心付近のわずかな領域から徐々に広がっていき、クムルスの細胞集団は常にこの拡大していくDelta の発現領域の縁に存在することが明らかとなった(図3)。つまり、背側を運命づける細胞集団の移動と尾部領域の拡大は時間的・空間的に一致した現象であった。ptc の機能を抑えた胚では、細胞集団の移動もDelta の発現も起こらない。このことから、クムルスの細胞集団は胚盤にだんだんとできあがる前後の位置情報を感知して、もしくは位置情報をつくる仕組みを利用して、中心から徐々に遠ざかるように動き、縁で止まるという仕組みがあるのではないかと考えた。
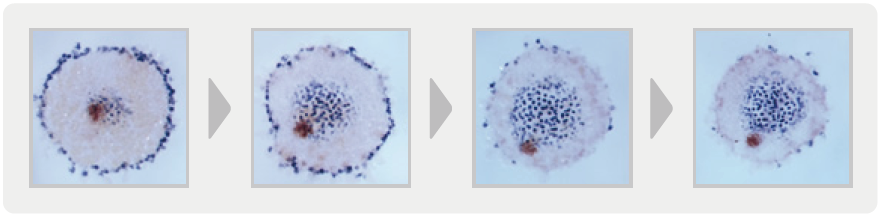
(図3)正常胚に見る尾の領域の拡大とクムルスの移動
受精後32時間(左)から36時間(右)まで。
胚盤の中心付近でDelta 遺伝子を発現する細胞は青色に、クムルスの細胞集団は茶色に染色した。
この考えを踏まえてもう一度、hh 遺伝子の機能を抑えた胚を解析した。当初の観察では、胚盤の前後のパターンの形成には明らかな異常が見つかっていたもののクムルスの動きは正常に見えた。しかし、hh 遺伝子、smo 遺伝子のそれぞれをはたらかなくした胚をビデオ撮影によってより精密に観察すると、クムルスが縁にたどり着いた後も止まらずに、縁に沿って動き続けるものが見つかった(図4中央)。クムルスが縁で止まっているか、動いているかという違いは、ビデオを通してはじめて区別できるものだった。hh やsmo の機能を抑えた胚では、Delta のように胚盤の中心で発現する遺伝子が縁まで拡大して発現し、縁で発現すべき遺伝子の発現が失われる。このため、クムルスは縁にたどり着いた後も自分が縁にいるという判断ができずに動き続けてしまうのではないかと考えることができる。
さらに、ビデオでの観察により、クムルスが止まらないもののほか、動かないものや遅れて動き出すものも見つかった(図4右)。この違いはクムルスが動き出す直前のDelta の発現領域の広がりに関係しているようだ。smo やhh のはたらきが強く抑えられると、Delta の発現は徐々に広い領域へと拡大するのではなく、胚盤全体で同時に始まってしまう。これとクムルスが動かないことに強い相関が見られた。このことから、背側を誘導するはたらきをもつクムルスの細胞集団が胚盤の中心から周縁へと動いていくには、前後の位置情報が徐々に形成されていくことが必要であると考えられる。
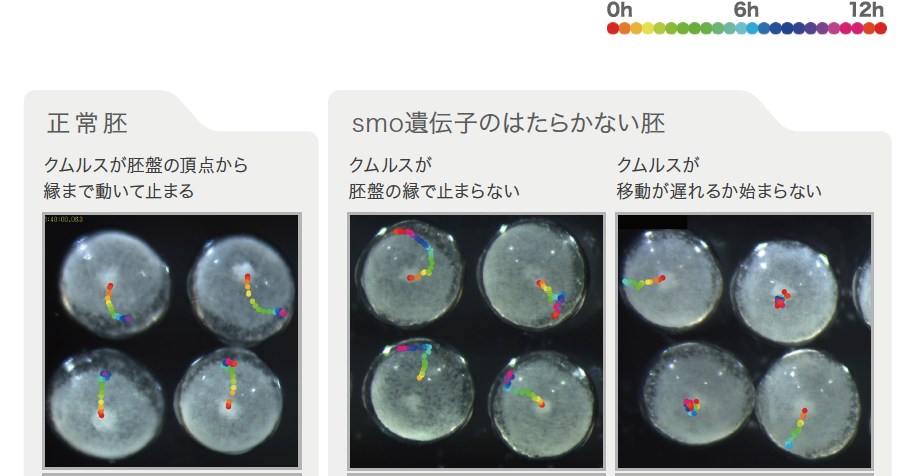
(図4) クムルスの動きに見られるさまざまな異常
4.前後のパターンを徐々につくる分子のせめぎ合い
次に、前後のパターンを胚盤の中心から周縁へ徐々につくる仕組みについて考えたい。これまで見てきたように尾部の形成に関わるDelta の発現は、正常胚では胚盤の中心付近から徐々に広い領域へと拡大していき、一方、hh やsmo のはたらきが強く抑えられた胚では胚盤全体から同時に始まる。実は、hh 遺伝子は胚盤の周縁で(より早い時期には胚盤の外側で)発現している(図5)。これらのことを総合すると、クムルスが動き出す以前には周縁で分泌されたHhが胚盤の中心付近まで拡散しPtcとSmoを介してDelta の発現を胚盤全体で抑えているのだが、次第にHhの拡散距離が短くなり、Hhが届かなくなった胚盤の中心付近から徐々にDelta の発現が始まると考えることができる。PtcにはHhの拡散を妨げる作用もあることが他の動物の研究で示されており、まだ仮説だが、このPtcの作用がHhの拡散距離の変化に関わっているようだ。先に述べたとおり、HhがPtcに結合すると、Smoが細胞内へとシグナルを伝達する(図6右)。さらに核内へとシグナルが伝達されると、頭部形成に関わる遺伝子などのいくつかの遺伝子に加えてptc 遺伝子の発現も促進される。つまりHhを分泌する細胞に近い細胞ほど細胞膜上のPtcが増え、そして増えたPtcはHh分子の拡散を妨げる。このようにして胚盤の中心から周縁へ徐々にパターンが形成されるのだろう。
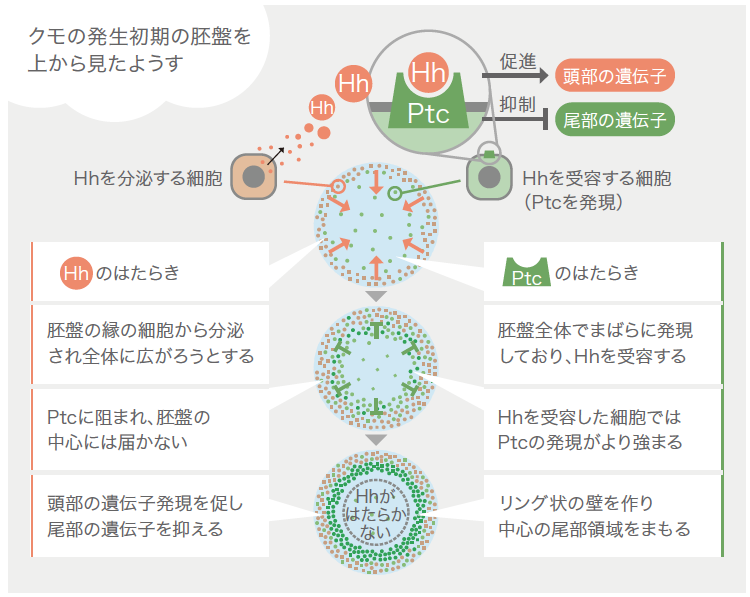
(図5)前後のパターンをつくる分子のせめぎ合い
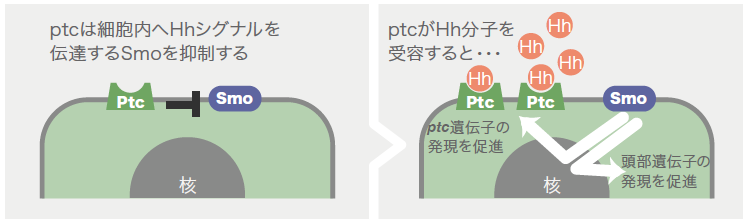
(図6) パッチトはHhシグナルを抑制する
5.放射相称のパターンの形成から二つの軸を直交させる
胚盤の中心から周縁へのパターン形成がクムルスの移動に必要であることが分かってきたが、クムルスの細胞集団が実際に分子レベルで何を感じているのかはまだ分からない。それは胚盤の中心から広がっていくものかもしれないし、中心から失われていくものかもしれない。いずれにしてもHhシグナルによって制御されるものであろう。今回の私たちの発見で非常に興味深いのは、前後軸と背腹軸の二つの軸が直交するという現象が、将来の前後軸につながる胚盤の放射相称のパターンを形成することで達成されている可能性があることだ。一つの軸の形成から直交する二つの軸をもつ体を形成することは、個体発生でも、動物進化でも、左右相称の体づくりの起源を思い起こさせるものである。今回明らかとなったPtcやSmoを含むHhシグナルの仕組みが、発生と進化をつなぐ新たな展開のきっかけとなれば面白い。
秋山-小田康子(あきやま・おだ・やすこ)
日本学術振興会特別研究員。1998年東京大学大学院理学系研究科博士課程修了。科学技術振興機構さきがけ研究21研究員、生命誌研究館奨励研究員などを経て、2009年より現職。2001年より生命誌研究館ハエとクモ、そしてヒトの祖先を知ろうラボにて研究。














