RESEARCH
食性転換は食わず嫌いをやめるところから
1.食性の進化を研究するためのモデル生物
現在世界中に生息し、遺伝学のモデル生物としてよく知られるキイロショウジョウバエも、もともとはアフリカ中央部に分布する “地域固有のショウジョウバエ” の一つに過ぎなかった。キイロショウジョウバエに最も近縁なグループは今もアフリカ大陸とその周辺に生息している。
ショウジョウバエはほとんどの種が、多様な植物の果実で繁殖する広食性だ。ところがアフリカ大陸東岸のインド洋に浮かぶセイシェル諸島の固有種セイシェルショウジョウバエは、ヤエヤマアオキ(註1)の果実だけで繁殖する単食性を示す。このような食性の違いはどのようにして進化したのだろうか。

(註1) ヤエヤマアオキ(Morinda citrifolia)
アカネ科ヤエヤマアオキ属の常緑高木。果実は強烈な悪臭を放つ。「タヒチアン・ノニ」の呼称で知られる健康飲料の原料でもある。
2.好き嫌いをきめる遺伝子を探す
セイシェルショウジョウバエは、キイロショウジョウバエとの共通祖先から約500万年前、最も近縁なオナジショウジョウバエからは40~200万年前に別れたと考えられている。布山喜章(東京都立大学教授:当時)らは、ヤエヤマアオキを好むセイシェルショウジョウバエとヤエヤマアオキを嫌うオナジショウジョウバエをかけ合わせた雑種個体はヤエヤマアオキを嫌うことを見出した。つまり、「好き」は「嫌い」に対して劣性に遺伝する形質なのだ。さらに高度な遺伝学の技が使えるキイロショウジョウバエとの間にも雑種を作り、この行動の違いが染色体上のごく限られた領域に起因することを突き止めた。ちょうどそのころ解読されたキイロショウジョウバエのゲノム情報を元に、該当する領域を探索した結果、2つの匂い物質結合タンパク質(OBP)遺伝子 Obp57d と Obp57e が、好き嫌いをきめる遺伝子の候補として浮かび上がってきた。
OBP(註2)は、昆虫が匂いや味などの化学物質を手がかりに食草を見分ける際の受容にはたらく分子である(図1)。キイロショウジョウバエはOBP遺伝子を全部で51種類持っているが、どうやらそのうちの2つがヤエヤマアオキの好き嫌いに関わっているらしい。
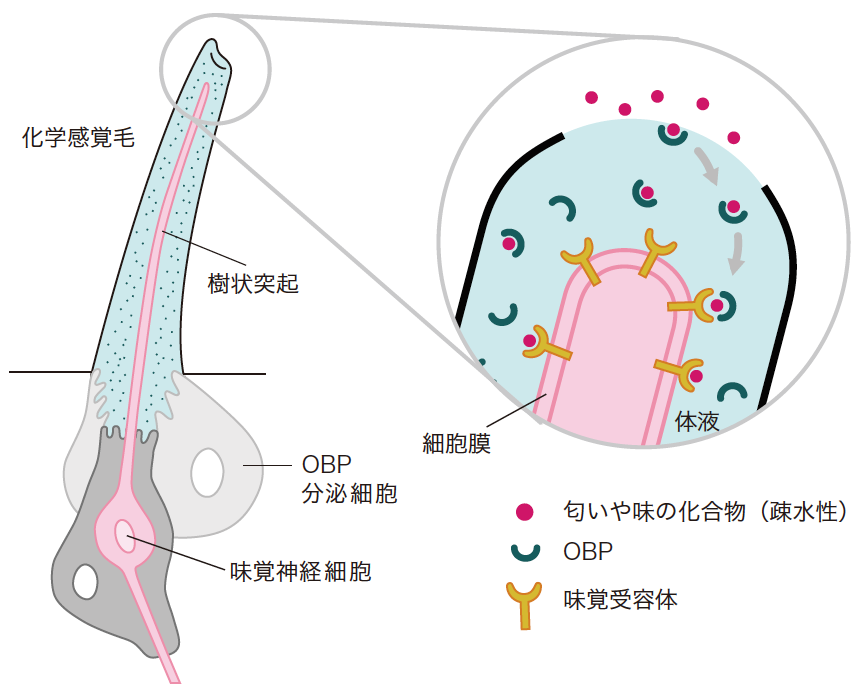
(図1) 嗅覚・味覚受容におけるOBPの役割
匂いや味の化合物は、化学感覚毛の表面に空いた小さな穴から毛内部に入り、神経細胞樹状突起の細胞膜に並ぶ受容体に結合して知覚される。化合物が疎水性の場合は、OBPが結合することではじめて体液に溶け込み受容体に辿り着くと考えられる。
(註2) 匂い物質結合タンパク質:OBP(odorant-binding protein)
昆虫の化学受容器の感覚細胞ではたらく一群の可溶性分泌タンパク質。
3.食わず嫌いを克服するきっかけ
2つのOBP遺伝子のはたらきを確かめるために、私たちはヤエヤマアオキに含まれる毒性のある脂肪酸、オクタン酸を用いた産卵場所選択試験(図2)を行った。意外なことに、ヤエヤマアオキを嫌うキイロショウジョウバエやオナジショウジョウバエも、オクタン酸の濃度が低い場合はそれを好むことがわかった。好き嫌いは決して二者択一の反応でなく、濃度があがるにつれて好きから嫌いへと変化するのである。ここでキイロショウジョウバエを遺伝子操作して Obp57d と Obp57e を取り除いたところ、ヤエヤマアオキに含まれるのと同じ濃度の比較的高濃度のオクタン酸入り培地も好むようになった。つまり、ショウジョウバエはもともと発酵した果実の目印として脂肪酸を好む性質を持っているが、その中で毒性を示すオクタン酸に対してだけは Obp57d と Obp57e のはたらきによって好みが抑えられ、嫌いになっているのである。
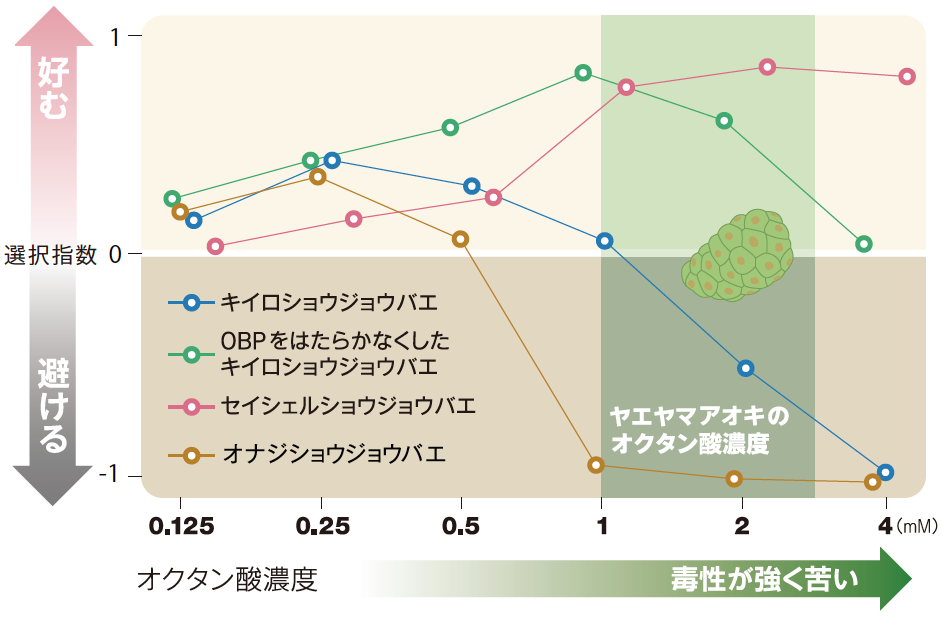
(図2) 産卵行動にともなう培地の選択
選択指数が1のときはすべての卵をオクタン酸入り培地に、-1のときはすべての卵を対照培地に産んだことを示す。キイロショウジョウバエやオナジショウジョウバエはオクタン酸濃度が低いと好む傾向があるが、濃度が上がるにつれて対照培地にしか産まなくなる。一方、セイシェルショウジョウバエは低濃度のオクタン酸の有無は区別できないが、濃度が上がるとオクタン酸入り培地に産むようになる。
キイロショウジョウバエやオナジショウジョウバエの Obp57d と Obp57e は脚にある苦味を感じる感覚毛で発現している(図3)。おそらく毒性のあるオクタン酸は苦味として受容されているのだろう。一方、セイシェルショウジョウバエの Obp57d と Obp57e は、遺伝子発現に必要なプロモーター配列が変化したために、遺伝子はあってもはたらかなくなっているということがわかった。
食べると危ないオクタン酸を、食べる前に察知する遺伝子 Obp57d と Obp57e は、食わず嫌い遺伝子と呼んでもよいだろう。実はセイシェルショジョウバエはオクタン酸の毒への耐性も進化させているが、食わず嫌いのままではいくらオクタン酸への耐性があってもヤエヤマアオキの果実に産卵することはない。セイシェルショウジョウバエの進化の過程における食性転換は、耐性獲得という生理的変化よりも、食わず嫌い遺伝子のはたらきを失うことによる行動的変化がきっかけとなって起こったと考えてよいのではないだろうか。
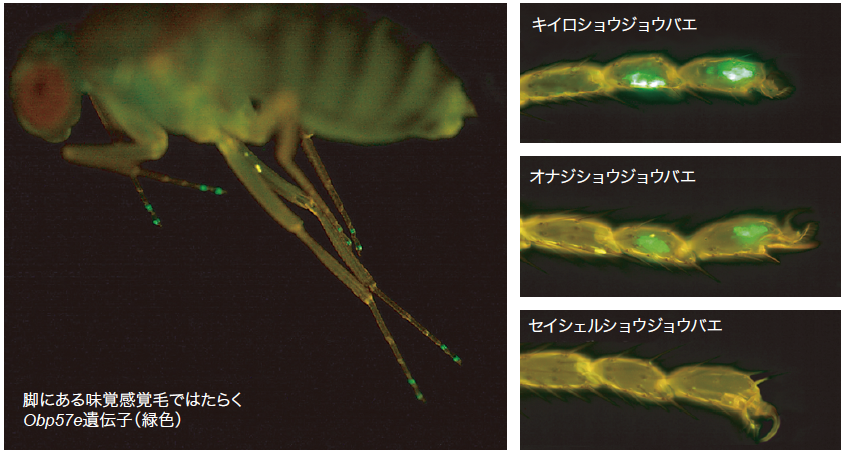
(図3) Obp57e 遺伝子の発現を見る
Obp57e 遺伝子の転写制御配列を蛍光タンパク質(GFP)遺伝子につないでキイロショウジョウバエに導入すると、各脚の先端にある化学感覚毛に付随する細胞でGFPの発現が見られた。これらの感覚毛は苦味を感じることができると報告されている。オナジショウジョウバエの転写制御配列を使った実験でも、同じ細胞でGFPの発現が見られたが、セイシェルショウジョウバエの転写制御配列はGFPの発現を誘導しなかった。
4.遺伝子から行動へとつながる味覚受容のしくみの進化
ヤエヤマアオキの好き嫌いのカギとなる遺伝子は Obp57d と Obp57e であることがわかったが、これらの遺伝子がどのように進化してきたかを知るために27種のショウジョウバエで Obp57d/e 遺伝子座のゲノム構造を比較解析した(図4)。2つの遺伝子は染色体上に隣り合わせて存在しており、元は一つの遺伝子だったようだ。遺伝子重複により Obp57d と Obp57e が生じ、いくつかの系統ではさらに数を増やしていったと考えられる。一方で、 Obp57d と Obp57e のどちらかが失われたり、たくさん増えた Obp57d のうちのいくつかが壊れてしまったりすることも頻繁に起きている。

(図4) Obp57d/e 遺伝子座の進化の様子
OBP遺伝子のアミノ酸配列と種間の系統関係を合わせて解析した結果。祖先種では一つだったOBP遺伝子(黄)から生じた Obp57d(青)と Obp57e(赤)はほとんどの種で維持されているが、いずれかを失った系統もある。機能分化を伴わない遺伝子重複は頻繁に起こるが、その後、D. takahashii と D. elegans で見られるように偽遺伝子化(遺伝子の構造が壊れること:d-pseudoと表示)してしまうようだ。
このようなOBP遺伝子の数の変化は、これらの種の食性と何かつながりがあるだろうか? この問いに答えるためには、それぞれのOBPの分子機能の違いから、それがもたらす味覚神経の応答の変化、さらには行動に表れる影響、と階層を横断する形で味覚受容のしくみの進化を明らかにしていく必要がある。
5.食わず嫌いできまる食性と好みできまる食性
セイシェルショウジョウバエのように、小さな島に隔離された環境で暮らす集団では、突然変異によって遺伝子が壊れても、自然選択を受けずにそのまま次世代へ伝わる確率が高い。実際、セイシェルショウジョウバエのゲノムではたくさんの偽遺伝子が生じている。このような “ゲノム崩壊” が、一方ではいわば進化の実験場となり、セイシェルショウジョウバエが毒のある「禁断の果実」を食べはじめる契機となったのは興味深い。
もともと広食性で、多様な植物の果実の中から食べてはいけないものだけを選別するための遺伝子を進化させてきたショジョウバエでは、食わず嫌い遺伝子を失うことが食性転換のきっかけになり得ることがわかった。一方、アゲハチョウをはじめとする、好みが細かく決まった狭食性や単食性の植食性昆虫では、目印となる化合物の組み合わせによって食草を識別することが知られている。しかも多くの場合、近縁種間でも好みが異なる。このような “好みに基づく食性” はどのようなしくみで決まり、また別の好みへと進化するのだろう。アゲハチョウとショウジョウバエ、それぞれの味覚受容のしくみを対比することで、食性と進化の関係の理解が深まることを期待している。
狭食性の鱗翅目の進化を探るのにナミアゲハに着目した。チョウが化学感覚毛で匂いや味を感じとれるのは、植物が分泌した化合物を神経細胞の受容体まで運ぶタンパク質があるからだ。その遺伝子を探すと、CSP(化学受容タンパク質)遺伝子19個とOBP遺伝子3個が見つかった。ゲノム構造を見ると、CSPは特定の染色体上で頻繁に遺伝子重複した形跡がみられることから、ナミアゲハの食草選択のカギはCSPの進化にあると期待している。食草を識別する化合物の組み合わせは、同じミカン科食性のアゲハチョウでも種間で異なる。現在その詳細を生体内の生理反応から解析する実験を進めている。(尾崎克久)
松尾隆嗣(まつお・たかし)
1997年東京大学大学院農学生命科学研究科中退、博士(農学)。豊橋技術科学大学助手を経て、1999年より東京都立大学理学研究科助手。現在、首都大学東京大学院理工学研究科助教。