RESEARCH
多様な細胞分裂様式に見る植物の進化
被子植物の細胞には、細胞骨格である微小管形成の中心となる「中心体」がありません。水中の藻類は持っている中心体を用いなくなった経緯をコケ植物で調べると、上陸に伴う新しい生き方の模索との関係が見えてきます。
1.細胞分裂様式の進化をコケに見る
陸上植物は、水中で暮らす藻類から進化したと考えられている。陸上植物は、藻類に比べて立体的で複雑な形態をもち、根、茎、葉など高度に分化した器官からなる。一方藻類は、鞭毛をもつ単細胞生物が中心であり、多細胞体の場合も、平面的な構造をしているものが多い(図1)。
陸上植物と藻類の細胞を比較してみると、藻類は、動物細胞と同じく細胞内に「中心体」と呼ばれる構造を持っているが、陸上植物は持っていない(図1)。中心体とは、細胞を顕微鏡で観察する時に見られる、星状の構造の中心部のことである。細胞の形態を支える微小管網や運動を制御する鞭毛など微小管を主体とする構築物は、この中心体を起点につくられる。さらに、生物が続いていく基本である細胞分裂では、紡錘体をつくる起点になり、染色体を正確に分配する大事な役割を果たす。
陸上植物の細胞は、このように重要な役割をもつ中心体が存在しないという構造をもちながら、藻類・動物と同じように紡錘体をつくり、染色体を分配できる(図1)。観察をすると、細胞分裂の進行に伴い、紡錘体の起点ともなる微小管形成中心が細胞内を移動し、細胞内のさまざまな場所から微小管がのびていることがわかる。つまり、陸上植物は進化の過程で中心体なしで正確に細胞分裂を行う新しい方法を獲得し、独特の紡錘体形成様式をもったのである。
このような陸上植物の細胞分裂様式が、どのようにして進化してきたかを明らかにするため、私たちは、陸上植物の中でも起源の古いコケ植物の細胞分裂に注目した。すると、コケの生活環(註1)の中で見られるさまざまな分裂様式から、細胞分裂の進化の試行錯誤が見えてきた。
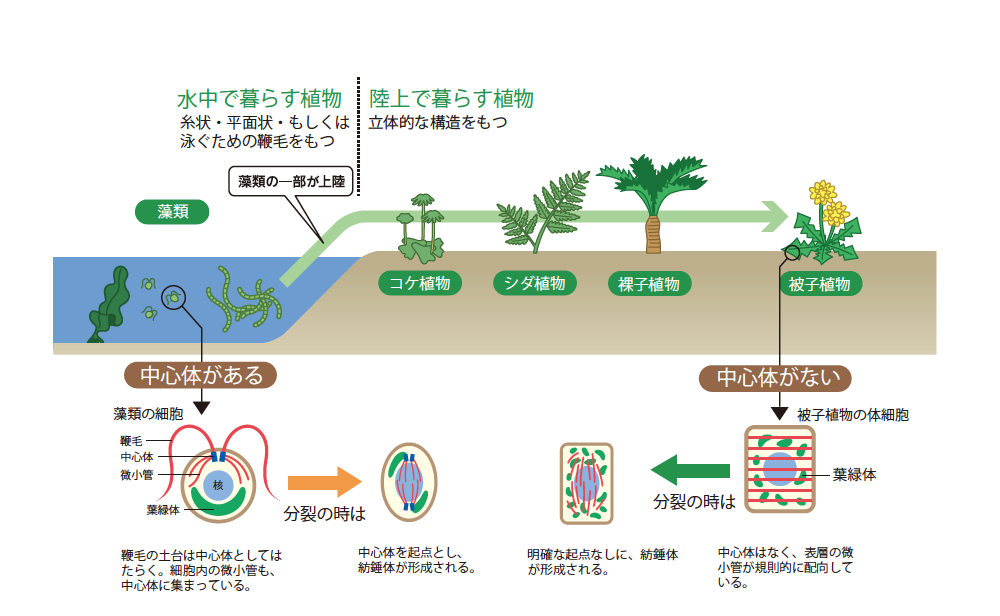
(図1) 藻類と被子植物の細胞分裂様式
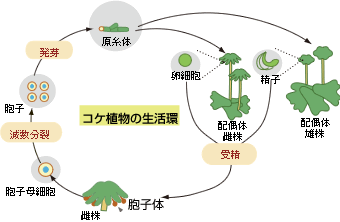
(註1) コケ植物の生活環(ゼニゴケ)
コケ植物は、配偶体世代と胞子体世代をくり返す。配偶体の雄株でつくられた精子は、雌株の卵細胞まで泳いでいき、受精する。受精卵から胞子体がつくられ、その中の胞子が飛び出して発芽することで、雌株と雄株からなる配偶体がつくられる。
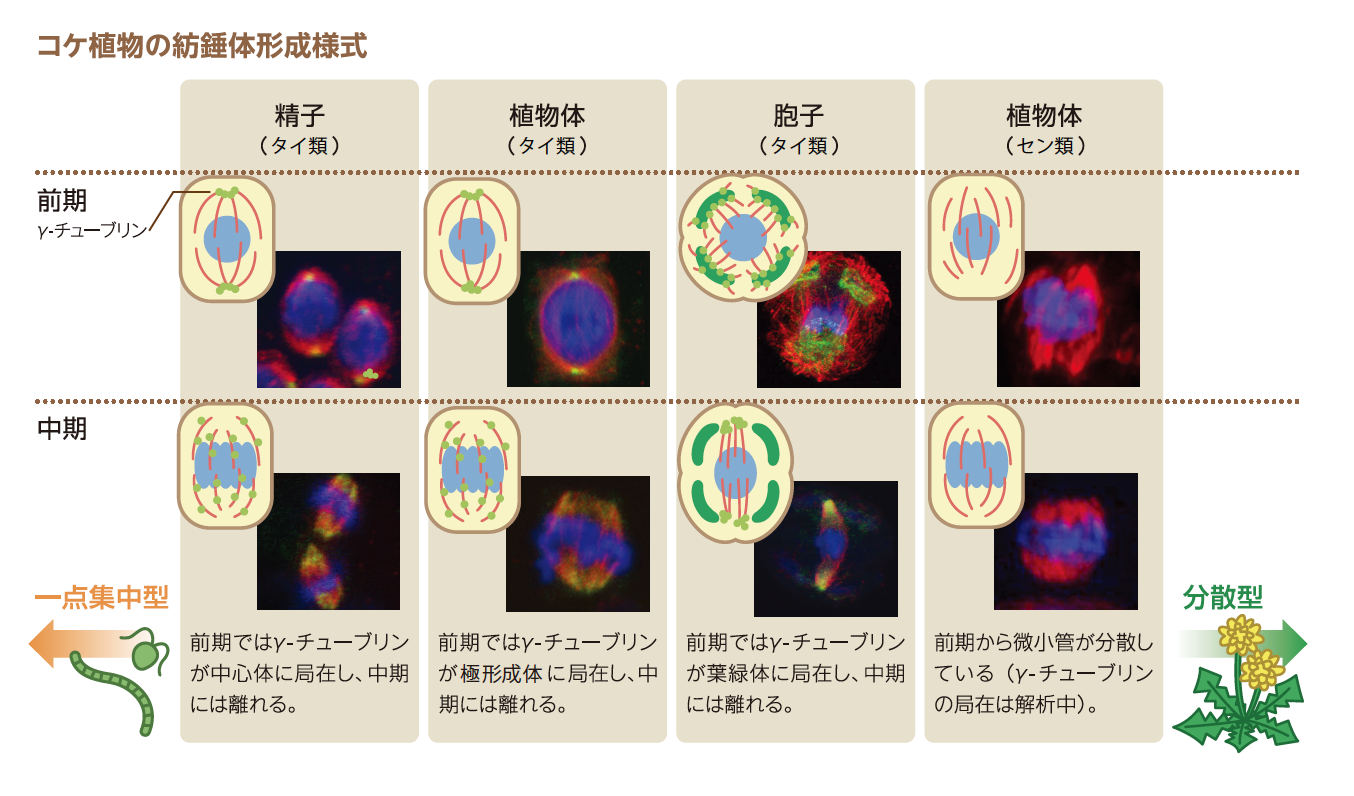
2.コケで見られる多様な紡錘体形成様式
コケ植物は、進化的に藻類と被子植物の中間段階と考えられており、紡錘体のでき方にもそれが見られる(表1)。例えば精子では、藻類と同じく鞭毛があり(写真1)、細胞内には中心体がある。精子をつくる時の細胞分裂では、中心体が微小管形成中心としてはたらき、紡錘体をつくり染色体を分配する。これは、コケ植物すべての精子で見られる。
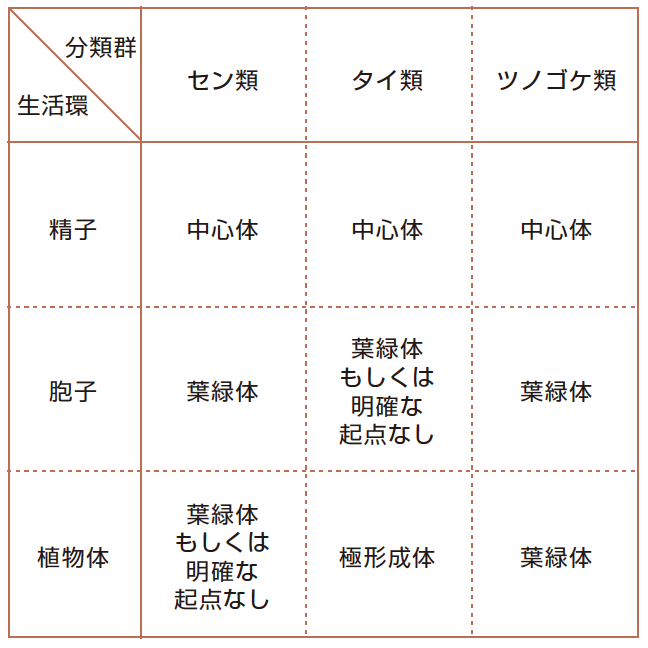
(表1) コケ植物に見られる微小管形成中心
精子形成では中心体から、胞子形成では葉緑体から、植物体では葉緑体・極形成体から紡錘体が形成される。セン類には、被子植物と同じように、明確な微小管形成中心が見られない。
また、ゼニゴケなどのタイ類(註2)の植物体における細胞分裂では、極形成体と呼ばれる中心体とよく似た構造が見られる。極形成体は中心体の内部にある中心小体は見られないが、中心体と同じように微小管形成中心としてはたらく。細胞内の一点から紡錘体がつくられるという点では藻類と似ているのである。
ところが胞子形成では、中心体も極形成体も存在しない。ここで、重要な役割を担うのが葉緑体だ。藻類やコケ植物でさかんに分裂している細胞は、基本的に1~2個の大きな葉緑体をもつ(写真2)。被子植物になると小さな葉緑体が常に多数見られるので、葉緑体を一つしかもたないことは、祖先的な形質であると考えている。ゼニゴケの一種、ケゼニゴケの胞子形成の減数分裂を見てみよう。葉緑体が一つしかない細胞では、それを娘細胞に確実に分け与える必要があるので、核の分裂に先立って葉緑体の分裂がおこるのだが(写真3)、その時、葉緑体表面が微小管形成中心となってそこから紡錘体がつくられ、葉緑体の分裂を追いかけるように核の分裂が行われるのだ。葉緑体が、動物細胞の中心体のように振る舞い、葉緑体がない細胞ができることを回避しているのである。
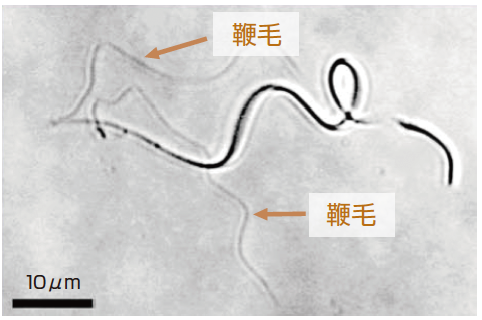
(写真1) ホソバミズゼニゴケの精子
二つの鞭毛が、頭部から後方にのびている。
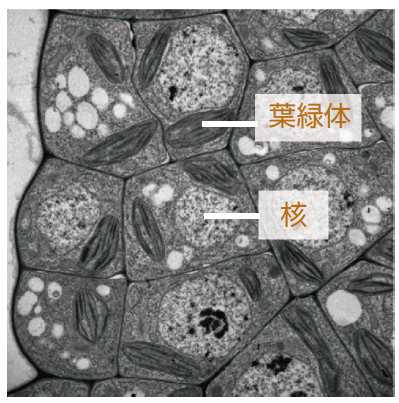
(写真2) 単色素体性のコケの体細胞
一つの細胞に、1~2個の葉緑体が存在する。
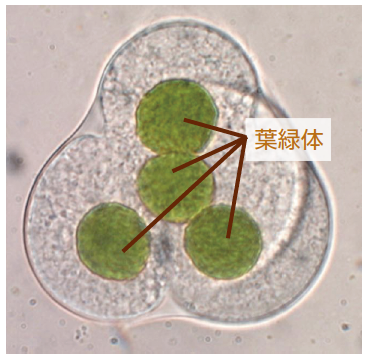
(写真3) ケゼニゴケの減数分裂
核分裂に先駆けて、葉緑体が分裂する。
さらに、葉緑体を多数もつセン類の植物体の細胞分裂では、被子植物と同様に明確な微小管形成中心がなく、紡錘体の極はタイ類のものほどはっきりしない。
コケ植物の紡錘体形成様式には、動物や藻類のように中心体を微小管形成中心とする一点集中型のものから、被子植物のように細胞質に分散した微小管形成中心を持つ分散型のものまで、さまざまな形態のものがみられることがわかった。これは、藻類から被子植物への進化の試行錯誤を反映していると考えてよかろう。
(註2) タイ類
タイ類は、ゼニゴケやジャゴケなどが含まれるグループをさす。コケ植物の中では、最もその起源が古いと考えられている。他のグループとして、ヒメツリガネゴケやスナゴケなどの蘚類、ツノゴケ類などが挙げられる。セン類が、被子植物の茎と葉を小さくしたような丈夫な茎葉体であるのに対し、タイ類は柔らかい茎葉体あるいは茎と葉の分化がない葉状体である。
3.多様な細胞分裂様式を司る共通の分子
中心体から微小管を形成する機能を直接担っているタンパク質の一つがγ-チューブリンであり、陸上植物も含めた真核生物全般に普遍的に存在する。動物細胞や藻類の中心体にはγ-チューブリンとそれに付随する分子でできたγ-チューブリンからなるリング状の複合体が存在し、そこが微小管形成中心となる(図2)。つまり、陸上植物の中で起源の古いコケ植物で、γ-チューブリンの局在をしらべることが、微小管形成中心の機能が中心体に集約された動物・藻類型から、細胞内に微小管が分散した被子植物型に変遷していく進化過程を考える上で重要なのである。
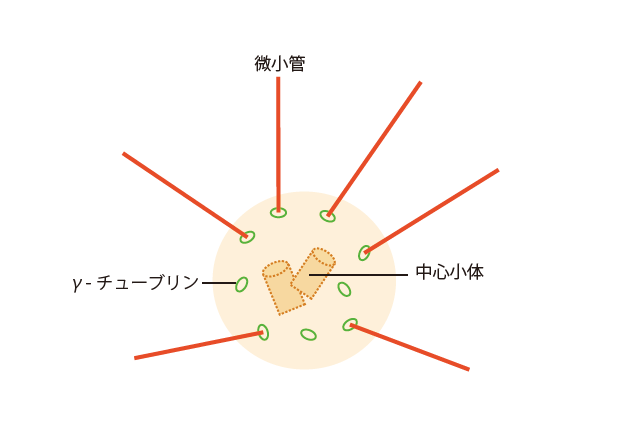
(図2) 中心体からのびる微小管
中心体上にあるγ-チューブリンを微小管形成中心として、α・βチューブリンが付加して、微小管がつくられていく。
そこで、コケ植物の生活環でみられる様々な細胞分裂様式においてγ-チューブリンの局在を調べてみた。精子形成過程の細胞分裂前期では、中心体がγ-チューブリンの抗体によって明瞭に染められる(図3)。中心体は最終的に精子の頭部に位置し、精子特有の微小管系や鞭毛の基部装置を形成する。タイ類の植物体で見られる極形成体にも、中心体と同様にγ-チューブリンが集中して局在する(図3)。
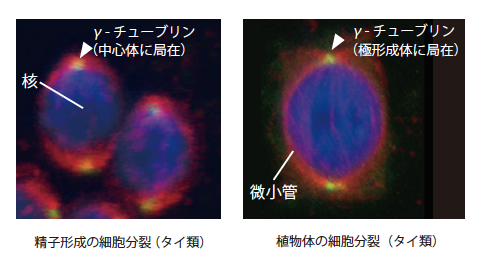
(図3) 中心体や極形成体を利用した紡錘体形成(分裂前期)
一方、葉緑体が微小管形成中心となる胞子形成では、4つの葉緑体から微小管が伸長する「四極微小管系」と呼ばれる構造が形成される。ここで私たちは、減数分裂の進行に伴い、γ-チューブリンの局在が、刻々と変化する様子を捉えることができた(図4)。減数分裂前期に葉緑体が2回分裂し、4個の娘葉緑体ができるのだが、分裂中の葉緑体から微小管が伸長し、このときまさにγ-チューブリンが葉緑体表面に局在する。その後、微小管が葉緑体から離れ、細胞の両極を起点とした紡錘体をつくる。この時、γ-チューブリンも葉緑体表面から離れ、紡錘体の極部分に集中するようになる。中期には微小管上に分散し、分裂後期になると、二つの娘核の間につくられた細胞板の前駆体である隔膜形成体(註3)と、娘核表面から生える放射微小管系(註4)にγ-チューブリンは局在する。このような減数分裂は2回行われ、2回目の分裂も、1回目と同様なγ-チューブリンの局在パターンを繰り返す。細胞分裂の進行によってめまぐるしく変化する微小管形成中心の位置とγ-チューブリンの局在が見事に一致するのである。コケ植物の葉緑体は、まさに中心体と同じはたらきをしていると考えてよいことになる。
ここから、コケ植物の様々な紡錘体形成様式は、藻類や動物と同じ分子機構をもったまま獲得されたことがわかる。細胞周期の進行に伴うγ-チューブリンの局在変化を司っている分子機構を、あらゆる生物に共通な機構として解明していくことは、今後の興味深い研究課題となる。
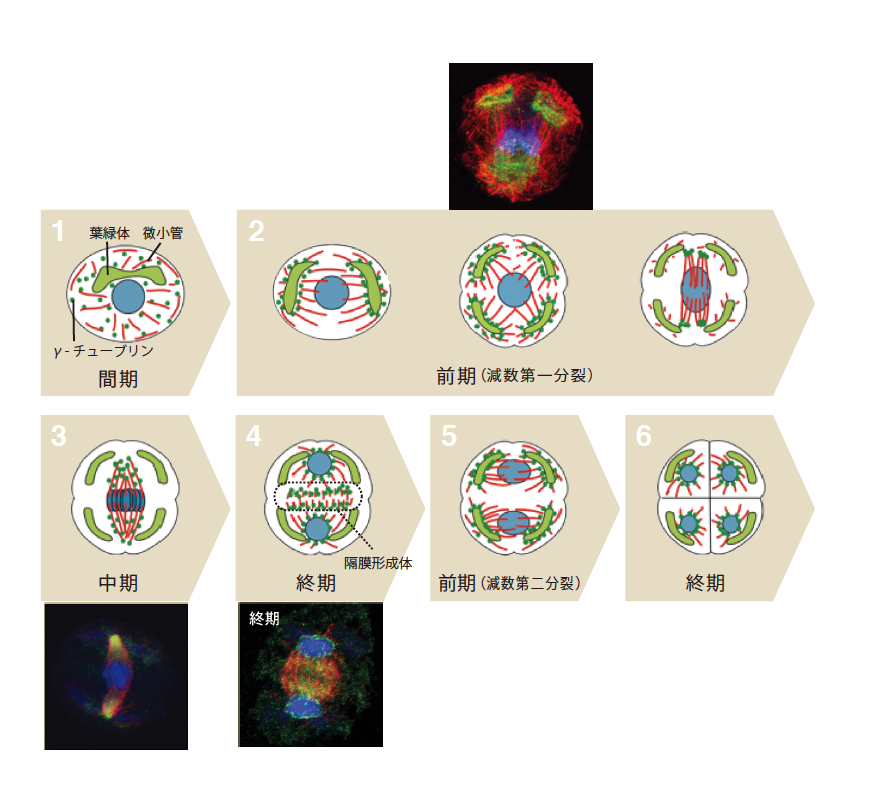
(図4) 胞子形成におけるγ-チューブリンの局在変化
核分裂に先立ち、葉緑体が4つに分裂し、4つの葉緑体にγ-チューブリンが局在する。γ-チューブリンは、葉緑体から細胞の両極(中期)、隔膜形成体(後期)へと移動し、4細胞に分裂後は、娘細胞の表面に局在する。
(註3) 隔膜形成体
細胞分裂終期に、分裂面を挟んで直交するように発達する微小管の集まり。ゴルジ体由来の小胞を細胞分裂面に集め、細胞板を形成しながらリング状に発達する。新しくつくられた細胞板は前期まで分裂準備微小管帯(コラム参照)が存在していた場所で親細胞の細胞壁に合着する。
(註4) 放射微小管系
娘核から放射状に微小管が伸びたもの。分裂準備微小管帯(コラム参照)がみられない細胞では、放射微小管系が隔膜形成体の形成位置を決定している。隔膜形成体は放射微小管系同士の境界でつくられる。これにより、娘細胞に均等に細胞質を配分している。
4.陸上植物の分裂様式の進化の系譜
コケ植物の紡錘体形成様式の多様性(図5)から、陸上植物の細胞分裂の進化の道筋を考えた。初期の陸上植物は、陸上に適応した体制を発達させる中で、鞭毛をもつ細胞を形成する時以外は、鞭毛の基部装置となる中心体を形成しなくなり、代わりに、葉緑体表面を紡錘体の微小管形成中心として使ったのだろう。また、1個の大きな葉緑体を持つ細胞から、多数の小さな葉緑体を持つ細胞へ進化していく過程で、微小管形成中心は葉緑体表面から細胞質に移行していったと考えられる。コケ植物の中には、複数の葉緑体をもち、紡錘体の極付近に葉緑体が集合するものがある。このような細胞では、葉緑体表面が紡錘体形成の起点となるという祖先的な機構がいまだに用いられているようにみえる。
コケ植物やシダ植物などの細胞分裂様式の多様性をさらに明らかにしていくことにより、陸上植物の細胞分裂様式の進化の系譜が探れるという方向が見えてきたので、これを更に確実なものにしていくつもりである。
(図5) コケ植物の紡錘体形成様式
コケの個体発生は系統進化をくり返す?
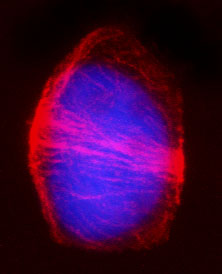
(写真4) タバコ(被子植物)の分裂準備微小管帯
細胞分裂前期の様子。赤く染まっている微小管が細胞の中央に集まっている。
例えばセン類は、胞子から植物体への発生過程で、糸状の藻類に似た体制(原糸体)を経るが(写真5)、その原糸体の細胞分裂では分裂準備微小管帯が形成されず、原糸体から立体的な体制をもつ植物体が発生する過程で初めて出現する。まさに、コケ植物の個体発生が系統進化的な歴史を繰り返しているようにみえるのである。陸上植物の複雑で立体的体制を生み出す細胞分裂面の制御は、分裂準備微小管帯の進化とともに獲得されたのだろう。
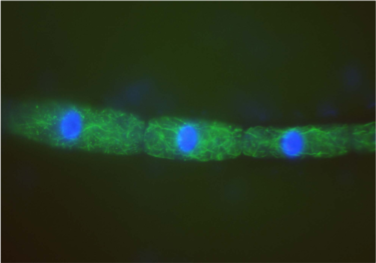
(写真5) セン類の原糸体
分裂した細胞が3つ連なっている様子。糸状の藻類のような体制をとる。緑色は細胞質内部で網目状に広がる微小管。
嶋村正樹(しまむら まさき)
2000年広島大学大学院理学系研究科博士課程修了、理学博士。日本学術振興会特別研究員、(財)地球環境産業技術研究機構研究員を経て2006年より広島大学大学院理学研究科助教。














