RESEARCH
生きものの多様性を支えるゲノムの水平伝播
種を特徴づけるゲノムを一つの細胞に納めるとどうなるだろうか。土壌で暮らす枯草菌に、水中で光合成をするシアノバクテリアのゲノムを丸ごと入れてみた。
枯草菌とシアノバクテリアのゲノムは、動物に例えるとクラゲとチョウほど離れている。それほど異なるゲノムが一つの細胞に納まったとき、細胞の中では何が起こるのか、全く異なる遺伝子ネットワークが一つの細胞内で長続きできるのか、新しい種としての変化が生まれるのだろうか、次々に生まれる問いを解析した。
1.DNAの垂直伝播と水平伝播
地球に暮らす生物は例外なく細胞からなり、その中に必ずゲノムが入っている。ゲノムとは生物がもつ遺伝情報の全体を意味し、DNAという物質として細胞の中に存在する。DNA中に並ぶATGCの4種類の塩基からなる配列がRNAへ写し取られ、タンパク質にまで翻訳されることで、生命活動が維持されている。セントラルドグマと呼ばれるこの生命維持のからくりは、塩基配列がいかに正確に子孫に伝えられるかにかかっている。しかし、正確さが過ぎると、激しい環境の変化に対応できず絶滅することになる。裏を返せばDNAが変化(註1)する柔軟さによって、遺伝情報の多様性が生まれ、多様な生きものの中から、その時々の環境に適応して、次世代にゲノムを伝えることのできた生物だけが結果として選択される。ダーウィンによって指摘されたこの原則は、目に見える動植物だけでなく、顕微鏡でしか観察できない微生物のレベルでも適用される。
DNAの変化は複製の誤りによって生じ、それが子孫に伝えられる。これは図1に示す縦の時間軸での変化であり、「DNAの垂直伝播」と呼ぶ。DNAの変化には、一塩基の変異だけではなく、遺伝子の重複、ゲノム内での転移、逆転も含まれる。これらが世代を経て徐々に蓄積し、種の多様性を生み出すのである。
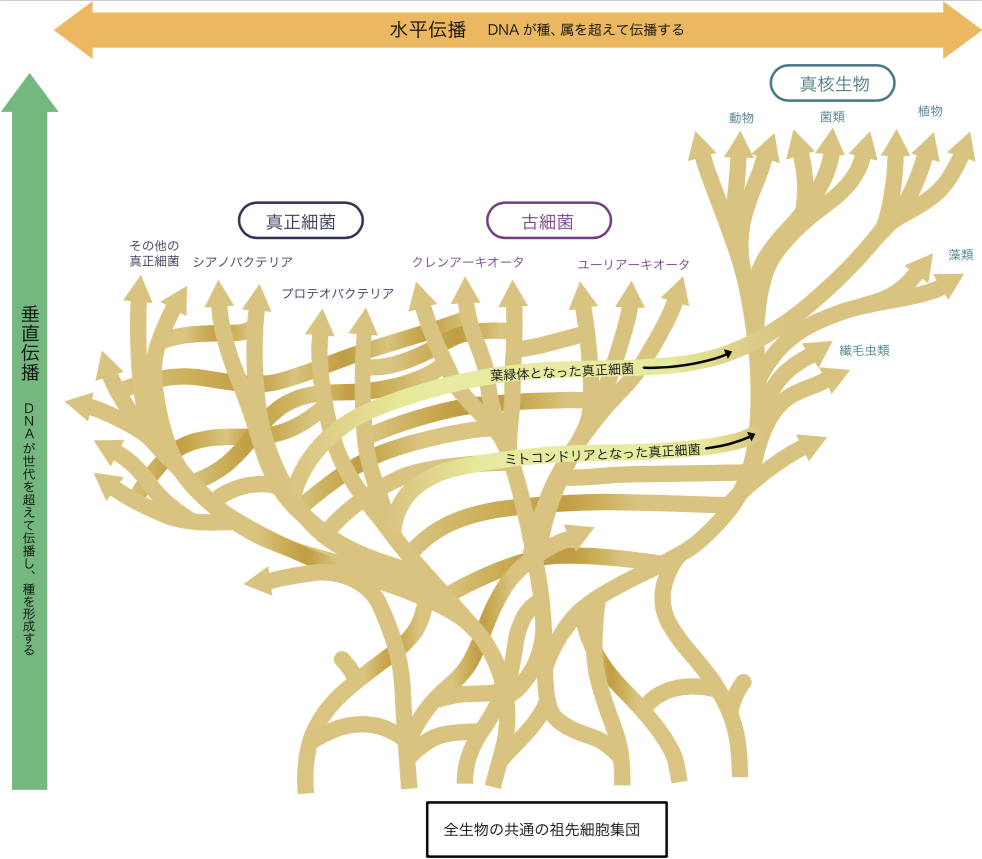
(図1) DNAの水平伝播
DNAは、世代を超えて伝播する(垂直伝播)以外に、種を超えて伝播(水平伝播)する。
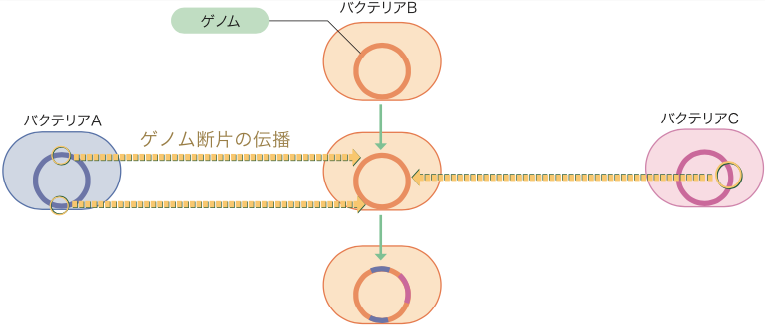
種ごとのDNAが世代から世代へと垂直に伝えられるのに対し、異なる種のDNAが侵入してゲノムに付加されたり、その一部が置き換わったりすることがある。これは「DNAの水平伝播」と呼び、図1の横軸に相当する。ゲノムDNAが核内に隔離されている真核生物では、ウイルス感染がその例だが、核を持たないバクテリアでは生物間で日常的に起こっている。こうしてゲノムに取り込まれたDNA断片は、そのまま子孫に伝達されるので、DNAの水平伝播はバクテリアの多様性を生みだす原動力なのである。現在私たちが見ているバクテリアゲノムは、種々の生物由来のDNAが取り込まれて、モザイク状になっていると考えてよい(図2)。
(図2) 全生物の進化系統樹
バクテリアのゲノムは異なる種のゲノムの断片が取り込まれて、モザイク状になっている。
ゲノムの全塩基配列を調べると、DNAの水平伝播は自然界での、以前予想されていた以上に頻繁に生じていることがわかってきた。一方、研究者は組み換えDNA技術を開発し、ありとあらゆる動植物、菌、ウイルスの遺伝子を大腸菌に人工的に水平伝播させている。クローニング(註2)と呼ばれる操作である。当初この操作は、自然界では起きないと考えられたが、その後に水平伝播の存在が明らかになってきたのだ。
(註1) DNAの変化
DNAの突然変異のこと。染色体、染色体の一部、遺伝子、ヌクレオチドにまで及び、変異の様式により、点突然変異、逆位、欠失、挿入、重複、転座などに分類される。
(註2) クローニング
特定のDNA配列を分離すること。遺伝子のクローニングは、ベクター(註3)に組み込んだDNA断片を宿主細菌(主に大腸菌)に導入し、目的のDNA断片を分離する。
2.実験室でゲノムの水平伝播を試みる
個々の遺伝子がクローニングできるなら、遺伝情報のすべてを担うゲノムを丸ごとクローニングすることもできるのではないのだろうか。実は、過去に自然界でゲノムが水平伝播したと考えられる名残りがある。細胞内器官のミトコンドリアと葉緑体であり、これらはそれぞれ独自のゲノムを持つ。ミトコンドリアはαプロテオバクテリア、葉緑体はシアノバクテリアが、過去に真核細胞に取り込まれたものなのだが(図1中央付近)、現在では多くの遺伝子が消失するか細胞の核ゲノムに移行している。つまり自然は、バクテリアゲノムを丸ごと取り込む(クローニング)手本を示していることになる。これに元気付けられ、私達も実験室でゲノムを丸ごと水平伝播させることを考えてみた。とはいえ、言うは簡単だが、技術的には大変困難な作業である。
困難な理由は二つある。まず、DNAという高分子は、溶液に溶かした状態で物理的な損傷を受けやすいことである。最も小さなバクテリア(マイコプラズマ Mycoplasma genitalium)のゲノムでも55万塩基対あり、500個ほどの遺伝子を持つ。これほど大きなDNAを無傷のまま、溶液で調製するのはとても困難だ。もう一つの理由は、クローニング原理そのものにある。現在主流のクローニング法では、大腸菌に特定の遺伝子を運び込むベクター(註3)として小さなプラスミドを用いており、運べるDNAのサイズに限りがある。ゲノムはその上限をはるかに超える大きさであるため、プラスミドでは運べない。
私たちはその方法論を徹底的に見直すところから始めた。大きなDNAを溶液で調製する問題は、小さなゲノム断片を一つひとつ、根気よく継ぎ足す方法をとることで回避できた。クローニングするDNAサイズの限界は、ゲノムを取り込む細菌を「大腸菌」ではなく「枯草菌」とし、枯草菌ゲノムそのものをベクターとして利用するという発想の転換によって解決した。枯草菌は、外部のDNAを積極的に取り込む独自の能力を持っており、巨大なDNAを安定してクローニングできる。
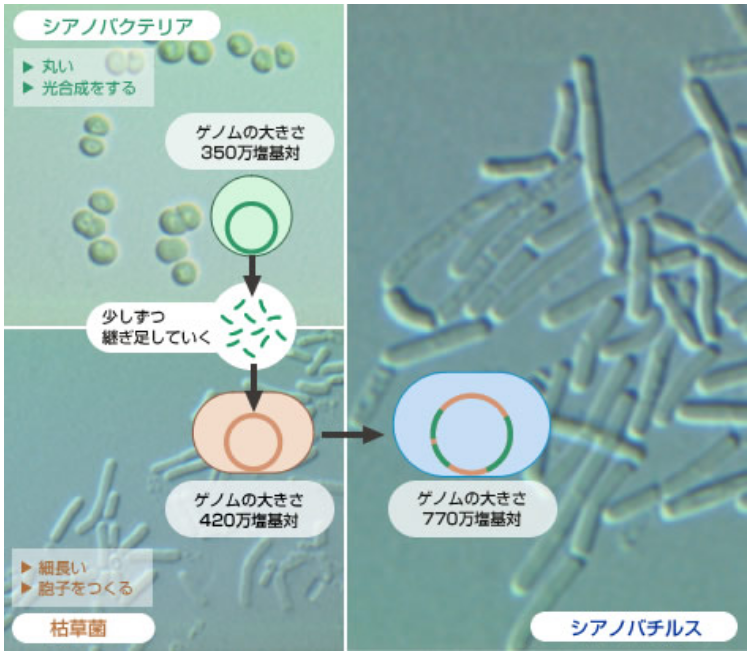
そうすると次なる課題は、枯草菌にどのバクテリアゲノムを取り込ませるかである。結論として、候補に挙げたのは、ゲノム解読が終了したばかりのシアノバクテリア(註4)を選んだ。そして、7年という歳月をかけて、350万 塩基対(遺伝子数は約3,200個)ものシアノバクテリアゲノムのすべてを枯草菌に移し、両種のゲノムが合体した「シアノバチルス」をつくることができた(図3)。ゲノムを丸ごとクローニングしたという成果は、類例がなく、かなりのインパクトを与えた(参考論文)。この成果の真に重要な点は、従来問われることの無かった数多くの課題を浮き彫りにし、検討を可能にしたことである。ここでは紙面の都合で二つの課題に絞りたい。一つは、種の同一性とゲノムとの関係であり、もう一つは、ゲノムを改変する具体的な手法に向けた模索である。
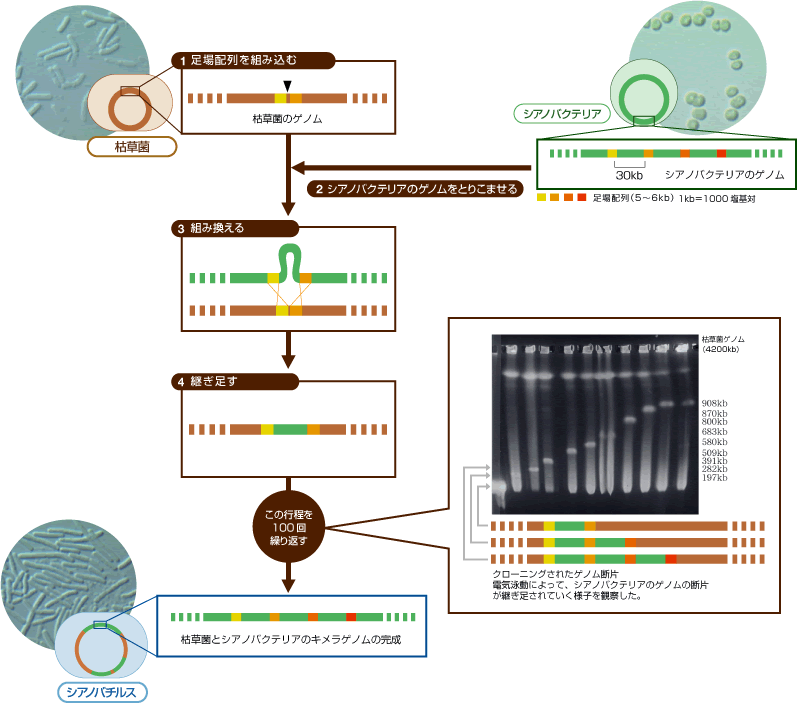
(図3) ゲノム丸ごとクローニングの方法
シアノバクテリアのDNA配列の中で「足場」となる配列を定め、それを枯草菌ゲノムにあらかじめ組み込み(1)、シアノバクテリアのゲノムを枯草菌に取り込ませる(2)。すると足場配列に挟まれたシアノバクテリアのDNA配列が相同組換えを起こす(3)ことで、そのDNA断片が継ぎ足される(4)。足場配列をずらしながら、この行程を100回くり返すと、350万塩基対のシアノバクテリアゲノムを枯草菌ゲノムにクローニングできる。
(註3) ベクター
組換えDNA技術において、外来性DNAを組み込み、宿主細胞中で増えることができるDNAを指す。自己複製能力をもつプラスミド、ファージなどを改良してつくられている。
(註4) シアノバクテリア
藍藻とも呼ばれる真正細菌の一種であり、光合成によって酸素をつくる特徴を持つ。1996年に全塩基配列が解読された。
3.シアノバチルスから種の同一性を考える
種の同一性とゲノムとの関係は、シアノバクテリアと枯草菌という異なる2つの種の遺伝子情報のすべてが一つの細胞に同居できるのかのという問いに集約される。実を言うと、ゲノムを丸ごとクローニングしていく過程で、最後まで同居ができない遺伝子があった。同居させると菌が生育できなくなってしまうのである。その遺伝子とは、メッセンジャーRNA(mRNA)からタンパク質を翻訳するリボソームRNAの遺伝子(rrn)であり、これが先に挙げた問いを解く鍵となる。
シアノバクテリアのrrnを欠くシアノバチルスには、両種のmRNAが存在するが、リボソームは枯草菌のものしかない。そして枯草菌リボソームは枯草菌のmRNAを優先的にタンパク質に翻訳しているようなのである(図4)。rrnを入れるとシアノバクテリアゲノム由来のタンパク質も合成されるようになり、その中に枯草菌に有害なものが存在するため、細胞が生き残れないのだろう。シアノバクテリアのrrnを欠くシアノバチルスの翻訳系は枯草菌の支配下にあり、シアノバチルスの形態も枯草菌より少し長細い形ではあるもが、光合成能もなく、基本的には枯草菌であると考えてよい。
一つの細胞内に異なる種のrrnが同居できない現象は、一つの種は一種類のリボソームしか持たないという経験則と一致する。この前提があるからこそ、私たちは、種の系譜を示す分子系統樹を描くことができる(図1)。とはいえ、異なる種のリボソームを含む生物は、本当に存在し得ないのだろうか。
この課題に取り組み、シアノバチルスをゆっくり生育する環境に移したら、シアノバクテリアのrrnを含む菌を得ることができた。その中には、形態的特徴として、枯草菌とは異なる丸や太い形の菌が見られた(図4)。生育環境の変化がシアノバチルスに何らかの影響を与えたに違いない。細胞の中でシアノバクテリアの翻訳系がはたらいているのか、さらには光合成をするようになるのか、今後詳細を確かめていきたい。
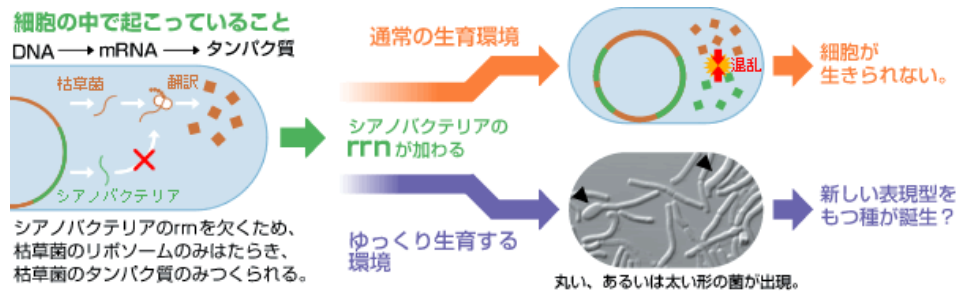
(図4) 翻訳システム(rrn)と環境によって決定されるシアノバチルスの生育
ゲノムすべてを移したところで、その情報が使われない限り、細胞の表現型は変わることはなく、種の同一性は保たれる。リボソームを中心とした翻訳系こそが、生育環境に応じて細胞システムを切り替え、種を決定する要因としてはたらくのだと考えている。
4.自然を手本にゲノムを改変する
ゲノムを丸ごとクローニングする第一の目的は、水平伝播によるゲノム進化を検証する系を創ることにあるが、これをバクテリアの育種技術に応用したいとも考えている。バクテリアは、私たちの生活に役立つさまざまな物質を生産する。バクテリアに有用な物質を効率的につくらせるには、遺伝子群を改変して目的の生合成経路を操作するというのが従来法である。そうではなく、新しい遺伝子を設計し、それを用いて生物を構築する合成生物学(註5)の概念と育種技術を融合させれば、ゲノムを一から設計し、有用な物質を生産するバクテリアをつくれるのではないかと期待できる。しかし、ゲノムを一から設計し、構築することは可能なのだろうか? 近年、DNAの化学合成技術が著しく進展して、PCR法(註6)や通常のクローニングで必須な鋳型DNAは不要になりつつあり、ゲノムサイズのDNAを化学合成だけで得られるようになった(註7)。つまり、ゲノムの設計図さえあれば、技術的にはどんな生物もつくることが可能になっているのだ。しかし私は、組み換えゲノムを自由に設計する前に、ゲノムとは一体何なのかという問いに向き合い、もう一度考え直す必要があると感じている。細胞の生育はゲノムだけでは成立しない。最初に述べたように、生育環境とゲノムとの組み合わせで選択されるのである。長い時間をかけて、その時々の環境に適応したゲノムを持つ生物だけが選択され、今を生きている。シアノバチルスの形態が生育環境によって変化したことは、ゲノムと環境との関わりの重要性を示しているのだと思う。この成果を出発点として、自然が構築したゲノムの青写真を理解したうえで、細胞膜を含むあらゆる生体構成要素の再点検へと進んでいきたい。
(註5) 合成生物学
構成的生物学とも言う。生命システムを構築することで、その理解を深める手段のこと。
(註6) PCR(polymerase chain reaction)法
DNAを増幅するための手法。増幅対象の鋳型DNAは、耐熱性のポリメラーゼによって指数関数的に増幅する。
(註7)
クレイグ・ベンダー研究所(米)の研究チームが、2008年にマイコプラズマ・ゲニタリウムのゲノム(58万塩基対)を化学合成のみで作成することに成功。
参考文献
Itaya, et al., Nature Methods 5, 41-43 (2008)
Itaya, et al., PNAS, 102, 15971-15976 (2005)
板谷光泰(いたや みつひろ)
1983年東京大学大学院理学系研究科博士課程修了、理学博士。米国NIH博士研究員、三菱化学生命科学研究所主任研究員を経て2007年より慶應義塾大学先端生命科学研究所教授。














