RESEARCH
「語る科学」
新しい形は「土台」から
-解剖学で見直す多様化のしくみ
研究マップ:最新のリサーチを、研究全体の中で位置づけよう。形態学の視点でまとめてみた。
1.付属肢:形態進化を考えるモデル
強靭なカニの鋏、鞭のようなエビの触角、爆発的な跳躍力を生むバッタの脚。形や働きは全く違う。それなのに、これらはみな、各体節に生じた一対の「付属肢」が変形したものであり、発生学的には相同器官である[図1](註1)。それだけではない。エビやカニの脚の根元には呼吸器官であるエラが付いているし、昆虫の翅は付属肢の一部からは生じたという説もあったりする。このように自由自在に形と機能を変え、挙げ句の果てに全く別の器官まで装備してしまう付属肢とはいったい何なのだろうか? 生物は進化の過程で様々な形態を産み出してきたが、節足動物の付属肢ほど多様化し、環境への適応に成功した例はない。付属肢は、形態進化(多様化)の仕組みを明らかにする格好のモデルなのである。
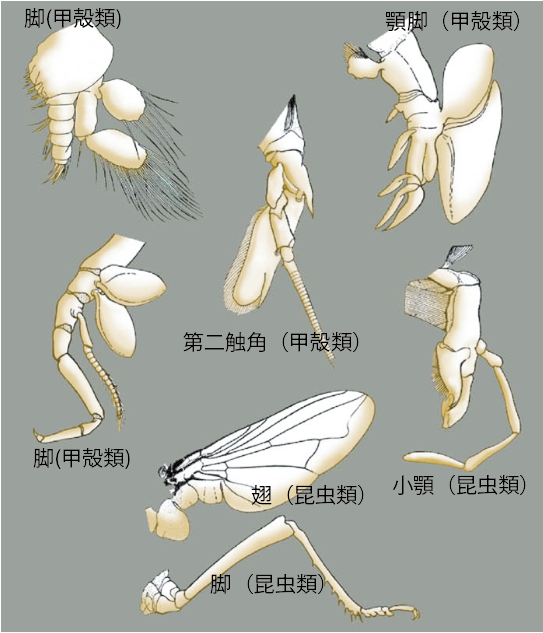
(図1)節足動物の付属肢いろいろ
節足動物の付属肢ほど形態的に多様化する形質はない。
(註1)発生学的には相同器官である
付属肢という用語はいくつかの動物群で用いられるが、ここでの付属肢は、各体節に1対あり、なおかつ明確な極性をもつものを対象としている。
2.付属肢の形態を理解するために
ところで、これだけ多種多様な付属肢のどの部分をどう解析していけば形態進化の仕組みが分かるのだろう? はっきりしているのは、そもそも付属肢とはどういう器官であるかを把握することから始めなければならないということである。しかし、極度に分化した付属肢を並べて眺めても、機能に適応した精巧なつくりにため息が出るばかり。しかも、一つの付属肢をとっても、大きさ、長さ、色、環節(註2)や刺の数、分肢の数や場所、筋肉の走行パターン、神経の分布、感覚器官の種類と位置、、、数えきれないほどの形質が混在していて、どの形質に着目すればよいのか見当もつかない。そこで、付属肢が本来どのような基本形から成り立っているのか、いわゆる「原型」(註3)のイメージをつくり、その原型に照らし合わせながら比較してみたらと考えることができる。 もしそれが可能なら、少なくとも、付属肢に見られる様々な形質がそれぞれ、どのように原型を変容させて獲得されたのかが分かるはずである。さらに複数の種間で比較をすれば、相互の相同性も把握できる。そこに系統関係の情報を加えれば、進化過程で原型がどのように変容してきたのか、その変容を通して発展してきた(あるいは維持されてきた)重要な形質が何であるのかを見出すことができる。原型を基準にすることによって得られるこのような理解こそが、形態進化の仕組みを明らかにする第一歩になるだろう。
(註2) 環節
節足動物の体幹および付属肢を構成する硬化した環状の節。硬い円筒をイメージしてもらえればよい。隣接する環節は膜質部分で連結され、環節間には一定方向への屈曲を可能にする関節が生じる。付属肢を構成する環節は肢節とも呼ばれる。
(註3) 「原型」
「原型」とは、頭の中でイメージする観念であり、存在の有無が問題にされる「祖先型」とは異なる。
3.原型を探る
付属肢の原型を見いだそうとする試みは1900年代前半から著名な古生物学者、形態学者らによってなされてきた。その中には、たとえば甲殻類にみられる二肢型付属肢(註4)を節足動物の付属肢の基本型とする考え方などがあるが、いずれの考えも節足動物にみられる多様な付属肢形態のすべては説明できず、今なお議論が残っている。では、少し範囲を広げ、節足動物以外の付属肢の形態を考慮に入れてみてはどうか? 前口動物群(註5)には節足動物の他にも、体節性をもち、なおかつ立派な付属肢を各体節に備えた動物群がある。環形動物に属する多毛類、いわゆるゴカイ類である[図2]。ゴカイ類の付属肢もまた、節足動物のそれに負けず劣らず多様化することで知られている。環形動物と節足動物が共通祖先に由来しているかどうかは議論の余地があるが、いずれにしても、節足動物の祖先を海産底生タイプの「環形動物らしきもの」であったと想定することは許されるだろう。そしてゴカイの付属肢は、体節性を獲得した生物がもち得る付属肢の比較的原始的なタイプと考えられる。そこで、付属肢の原型の探索を、ゴカイの付属肢と最も原始的な真正昆虫類(無翅外顎類)であるイシノミ類(註6)の付属肢との比較から始めた。

(図2) ゴカイ類とイシノミ類
イシイソゴカイ(右)とヒトツモンイシノミ(左)。
下はそれぞれの付属肢の電子顕微鏡写真。
ゴカイの付属肢は無関節肢で多分岐するが、イシノミの付属肢は関節をもつ環節肢である。
(註4) 二肢型付属肢
二叉型肢ともいう。基部側の肢節より、それぞれ数節からなる外肢、内肢を発するタイプの付属肢。甲殻類の下等なものや発生初期では、外肢と内肢の形態が類似することなどから、この二分岐した状態を節足動物の付属肢の基本形態を考える場合が多い。この場合、昆虫類の歩脚は内肢が発達したものと考えられている。
(註5) 前口動物群
先口動物、旧口動物ともいう。発生時に原腸陷入によって生じた原口がそのまま成体の口になる動物群の総称。節足動物、環形動物、軟体動物などが含まれる。ちなみに私たち脊椎動物は原口が成体の肛門になり、口は後に新たに開口する後口動物(新口動物)である。
(註6)最も原始的な真正昆虫類
(無翅外顎類)であるイシノミ類
イシノミ類は、口器の大顎が頭蓋と1か所だけ接続すること、翅をいまだ獲得していないこと、腹部にも付属肢が発達すること、成虫になっても脱皮を繰り返すことなど、高等昆虫類にはみられない原始的な形質が多数みられることから現存する真正昆虫類(外顎類)の中でもっとも原始的と考えられている。
4.環形動物の付属肢と原始昆虫類の付属肢
図2にあるように、ゴカイ類とイシノミ類の付属肢の外部形態は似ても似つかない。原始的昆虫類であるイシノミ類の付属肢は、節足動物の名のとおり関節をもった環節肢である。唯一、他の昆虫類と異なる特徴は、胸部第2、3節の付属肢の基節に甲殻類で見られるような外突起が生じることである。これに対し、ゴカイの付属肢は葉状で無関節であり、先端部で多分岐する。多毛類という名からもわかるように、2束のブラシのような剛毛が突き出しており、これによって砂利をかき分けて進むことができる。
通常、節足動物間の形態比較の場合は、関節構造(註2)を目印として対応関係を見いだしてゆくが、無関節のゴカイ付属肢が相手では到底無理である。では、どうするか?遺伝子の発現パターンを比較すればいいという意見もある。しかし遺伝子は発生過程の様々な部位で繰り返し機能するので、遺伝子発現パターンは形態を理解するうえでは決して万能ではなく、一つの形質にすぎない。つまり遺伝子発現の類似には、必ず形態学的な裏付けを加えなければ意味がなく、時として大きな判断ミスをおかす危険がある。そこで、まずは形態比較を徹底的に行い、注目する形質を絞り込むのがよいのではないか。形態は外部形態に限ったものではなく、神経系、筋肉系、循環器系などの内部形態もまた重要な比較形質である。なかでも能動的に動く器官の場合、筋肉の配置パターンがその動きをコントロールする重要な構造となる。付属肢も例外ではない。そこで、全く外部形態の異なるこの2つの付属肢を、筋肉系を通して比較してみた。
5.筋肉系の比較から見えてきたもの
解剖とスケッチを繰り返す。連続組織切片を作成して立体構築する。筋肉を特異的に染め出す。系統関係を考慮して二次的に失われているであろう筋肉を復元する。それらの情報を統合し図にする。そうして出来上がったそれぞれの筋肉系の模式図を並べてみた[図3]。見えてきたのは、外部形態の違いからは想像しえない類似性であった。ゴカイ類の付属肢筋肉として同定したものから、ゴカイ類に特有の2束の剛毛を動かすためだけの筋肉をすべてのぞいてゆくと、イシノミ類の付属肢の基節までの筋肉系ときわめてよく似た走行パターンが残る。ゴカイ類の付属肢は、あたかも原始的昆虫類の基節までの部分が発達し、いくつかの分岐を備えた形のようである。このような類似性は収斂の結果なのか、それとも共通祖先が存在していたことを示すのか、現時点ではわからないが、ある2本の筋肉の存在から付属肢の原型を思いつくヒントを得た。
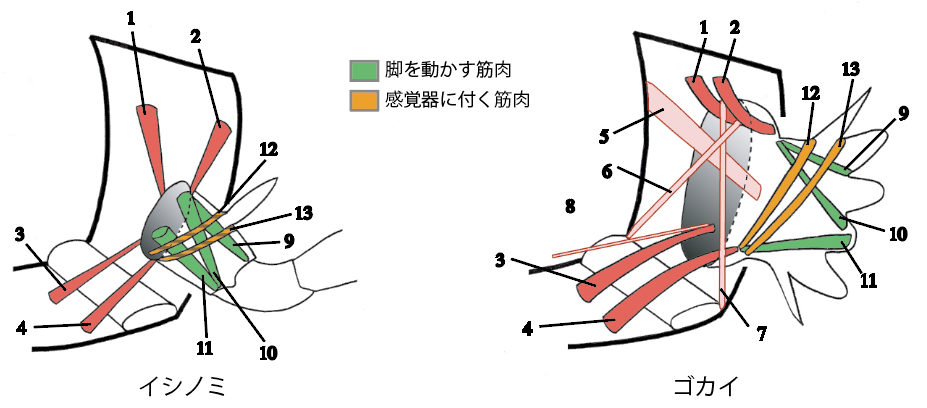
(図3) 筋肉系の類似
外部形態の顕著な違いに反して、筋肉系はよく似る。イシノミ類の付属肢で確認、および推測された筋肉(右)のすべては、同様のものがゴカイ付属肢の筋肉系(左)で見いだせる。ゴカイ付属肢の図では、剛毛を動かすための筋肉は省略してある。類似する筋肉には同じ番号をふっている。イシノミ付属肢の12、13番筋肉は推測されたもの。
6.背部感覚器官
ゴカイ類のたくさんに分岐した付属肢のうち、背側と腹側にもっとも近い位置にある2つの突起は、感覚器官としてはたらく。この突起は走査型電子顕微鏡で拡大しても区別はできない程、形態と機能が酷似している。それにもかかわらず、なぜか背側の感覚器にだけ2本の筋肉が付いているのである[図4]。一方、イシノミ類の付属肢の基節の背面側にある外突起は、オスとメスが交尾ダンスをするときに接触刺激を感じとる感覚器官としてはたらくとされている。現存するイシノミ類ではこの感覚器官に筋肉は付いていないのだが、この感覚器官の基部には単純な関節構造が残っていることや、甲殻類の相同な形質には2本の筋肉が付着いていることを考えると、イシノミ類の外突起(感覚器官)にも本来は2本の筋肉が付着していたと推測できる。すなわち、ゴカイ類とイシノミ類では、付属肢のタイプが全く違うにもかかわらず、いずれも背部に2本の筋肉でコントロールされる感覚器官を生じさせた可能性が高いのである。

(図4) オレンジ色で染めだされたゴカイ付属肢の筋肉
背側の感覚器官には2本の筋肉(矢印)が付着する。腹側の感覚器官には筋肉はつかない。
7.付属肢の原型と付属肢進化の新しいシナリオ
筋肉でコントロールされる感覚器官が、付属肢の背部に共通して存在する理由は今のところわからない。しかし、昆虫類の系統を辿ってゆくと興味深い事実に気付く。昆虫類で最初に翅を獲得したとされるカゲロウ類では、「翅」に加え、もう一つ特徴的な「気管鰓」という器官が発達する[図5-1]。これは水棲である幼虫時代に腹部体節の背面部側方に発達する呼吸器官で、付属肢と同様に1体節あたり1対生じる。実は飛翔器官である翅と、呼吸器官である気管鰓は、いずれも付属肢の一部から生じた形質であると考えられている。しかも、これらの器官はいずれも2本の直接筋によって動作がコントロールされているのである[図5-2]。もし、この直接筋がゴカイ類やイシノミ類でみられた背部感覚器官を制御する2本の筋肉と相同であるとしたら? こう仮定すると、形態の変化部位を偶然の結果とするこれまでの進化イメージとは違う、新しい付属肢進化のシナリオが出来上がる。すなわち、体節性をもった生物が付属肢を獲得した当初、すでに形態的な変化を引き起こせる土台が付属肢の背部領域に用意されていた。昆虫類の系統では、その背部領域を積極的に利用することで形態的な多様化が急速に進み、呼吸器官や飛翔器官など、それまでにはなかった新たな形質を生み出すことができた、というものである。もちろんこの場合、付属肢の原型は、「昆虫類の付属肢でいうところの基節までの構造を基本とし、その背部領域には、自律的な制御が可能な器官を生み出す土台があらかじめ備わっている」と考える。付属肢形態の革新的な進化とは、結局は、原型の中に用意された土台(註7)を上手く応用できるかどうかなのではないだろうか。
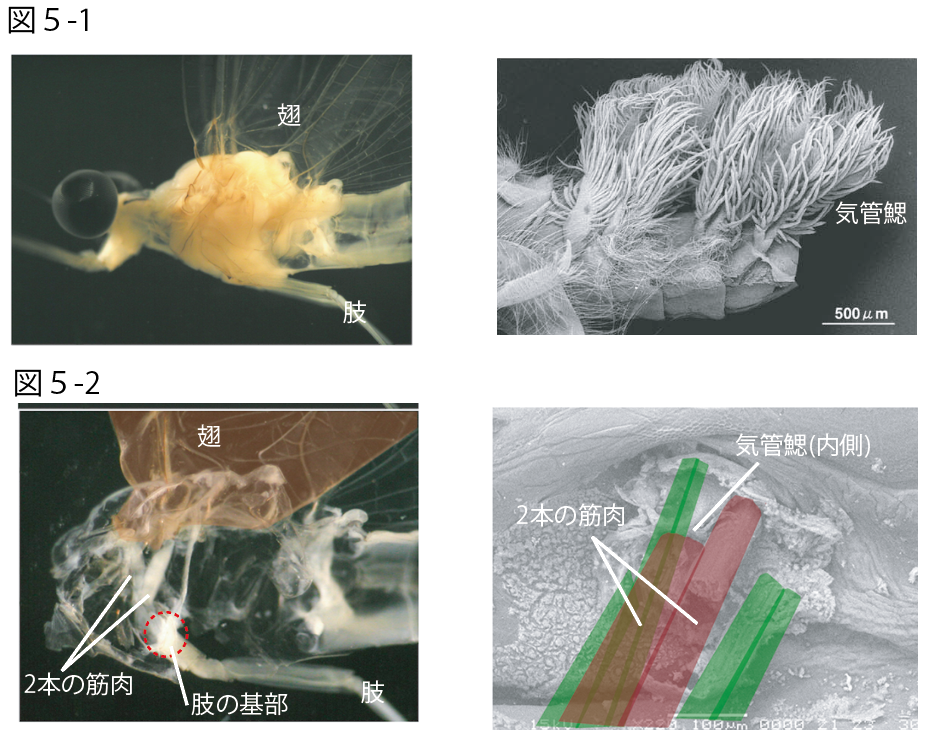
カゲロウの翅と気管鰓
(図5-1)外部形態
フタスジモンカゲロウの成虫の翅(左)と、幼虫に見られる気管鰓(右)。
(図5-2) 内部形態
成虫の翅に付く直接筋は肢の基部に2本でつながる(左)。幼虫の気管鰓に入りこむ2本の直接筋は赤色で、周辺の表皮に付着する筋肉は緑色で示す(右)。
8.核心へ
では、付属肢背部に想定した「土台」とはいったい何なのだろう。またはどういう状態にあることをいうのだろう? あるいは、なぜこの部分で形態的変化を生じさせることができるのだろうか? まさに「土台」の具体的な解明こそが、次のステップであり、この研究の核心である。我々はすでに、この土台に相当する領域が、発生過程では他の付属肢領域とは異なる細胞集団で構成されるという組織学的な情報を得ている。さらには、コオロギの付属肢では相当する領域に、オーガナイザー活性をもつwingless遺伝子が特異的に発現することも報告した(Niwa et.al. 2000)。これらの情報を元に、付属肢背部に備わった「土台」の実態を細胞や遺伝子のレベルで説明できるはずである。そして、実際にその土台が使われる(形態的変化が生じる)ための必要条件を明らかにできた時にはじめて、形態進化の仕組みが「少し」分かったと言えるのだろう。そこに向かって実験を進めていくつもりだ。
<謝辞>
本研究は開始当初から形態形成シグナル研究グループの林茂生グループディレクター、秋元愛技術員をはじめとする多くの方々から惜しみない支援をいただいています。特にイシノミ類については町田龍一郎先生(筑波大学大学院生命環境科学研究科/菅平高原実験センター)、カゲロウ類については東城幸治先生(信州大学理学部生命科学科)、ゴカイ類については高橋理氏(香川県)、三浦知之先生(宮崎大学農学部生物環境科学科)、西栄二郎先生(横浜国立大学教育人間科学部付属理科教育実習施設)より貴重なサンプルや各生物に関する情報を提供していただきました。ここに深く感謝いたします。
丹羽尚
独立行政法人理化学研究所 発生再生科学総合研究センター 形態形成シグナル研究グループ 研究員
1998年徳島大学工学研究科博士課程修了。国立遺伝学研究所 研究員、理化学研究所 基礎特別研究員を経て、2004年より現職。