RESEARCH
「語る科学」
なぜ今、クモなのか?
胚発生が描く進化の道すじ
1.動物の祖先はどんな形だったのか?
動物の進化、とりわけ私たち人間もその仲間である脊椎動物の祖先に関しては100年以上も前から研究者の間で熱い議論が交わされてきた。ミミズの仲間が脊椎動物に近い祖先だと考えられたこともあったし、クモ類がその候補に挙げられたこともあった。近年は、形態や化石を見る従来の研究に加え、分子系統学(DNA塩基配列の置換の蓄積を見て系統樹を描く方法)に基づく系統樹が描かれ、進化の大筋は分かったかのように言われている。だが、発生のしくみに関する遺伝学的な知識が増えれば増えるほど、動物の形の進化と系統関係を矛盾なく説明することが難しくなっている。とくに、系統的に離れているショウジョウバエ(節足動物-昆虫)と脊椎動物で、形を決める遺伝子のはたらきにさまざまな類似点があることが明らかになってきた。
とりわけ注目すべき2つの類似点を挙げよう。1つはHox遺伝子クラスターによる前後軸に沿った領域の特異化、もう1つはSog/Dpp遺伝子による背腹軸の決定である。体の形の決定の基本ともいえる前後軸、背腹軸(*註1)の形成に、ハエとマウスで共通の遺伝子がはたらいているのである。De RobertisとSasaiは、この類似は進化的保存によるものだと主張し、ハエと脊椎動物の共通祖先として仮想の動物Urbilateria(ウルバイラテリア)を提案した【図1】。Urbilateriaは、分子系統学による系統関係と合わせて、昆虫と脊椎動物の共通祖先としてだけでなくすべての左右相称動物の共通祖先として考えられた。Urbilateriaの仮説はこの分野の研究者に多くの影響を与え、現在でも強い支持を集めている。しかし、左右相称動物の共通祖先はもっと単純な形をしていたのではないかという考え方も根強い。左右相称動物の仲間であるウニ(棘皮動物)やホヤ(尾索動物)などの発生を見ると、共通な祖先は単純な形をしていただろうという考え方も充分有り得る。ハエと脊椎動物の類似が、Urbilateria仮説が主張するように本当に進化的保存によるものなのか、それとも単純な祖先から独立に進化して二次的に似たものなのか、左右相称動物全体を見なければ解決されない問題である。
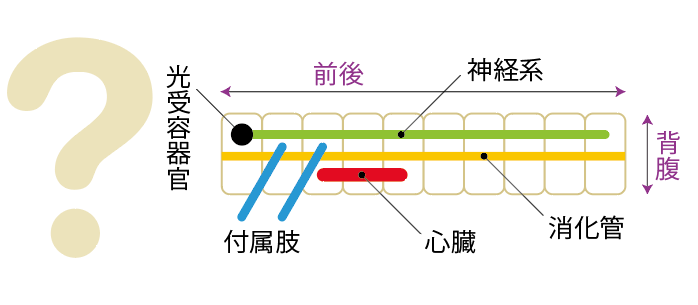
(図1)ウルバイラテリア
左右相称動物の共通祖先として提案されたUrbilteria。だが、未解決の問題も多い。
私たちは、ハエと脊椎動物の類似を本質的に理解するためには、発生生物学の見地から、次の2つの問題を解決する必要があると考えている。1つは、体の繰り返し構造(体節)に関してである。ハエでは外胚葉(体の外側)が、脊椎動物では中胚葉(体の内側)が体節形成の場となっているのはどうしてか? 第2は、脊椎動物の背腹軸形成では中胚葉が中心的な役割を果たすのに対して、ハエの中胚葉は背腹軸形成に直接関与していないのはどうしてか?いずれも体の形づくりの基本として重要な問題である。この問題を解くための糸口を、節足動物の一つであるクモでつかみたいと考えたのである。
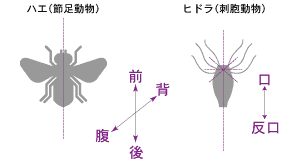
(註1) 前後軸・背腹軸
左右相称動物は、卵から幼体になるまでの胚発生の時期に、まずは前後・背腹の軸が決定し、それにしたがって体の節や器官がつくられる。相称とは、一つの線または平面の両側で動物の体の部分が互いに一致することをいう。ほとんどの動物は前後(頭部と尾部)、背腹は非対称性であり、左右相称を示す。より原始的な体の型には、原生生物やカイメンなどの非相称、クラゲやヒドラなどの放射相称がある。
2.なぜ今、クモなのか
多様な動物は体制の類似性に基づいて「門」にグループ分けされているが、前出のハエとマウスの発生を比べる場合、節足動物と脊椎動物という「門」を越えた比較となる。このとき、両者で体のどの部分がどの部分と比較できるのかという最も基本的な問題が生じてしまう。同じ動物門の中の比較では、この問題はある程度避けられる。そこで、それぞれの動物門の祖先を理解しようとする努力の中に新たな展開が期待できるのではないかと考えた。
研究対象としてクモを選択した理由のひとつは、ハエとクモの比較を通して節足動物門の初期の状態を理解しようと考えたことである。ショウジョウバエは分子遺伝学者の努力によって既に莫大な情報が蓄積し、「辞書」のような存在になっている。また、現在の分子系統学に基づくと、ハエとクモの最後の共通祖先は、現存のすべての節足動物の最後の共通祖先と一致する。化石記録上では、クモの属する鋏角類は昆虫類よりも1~2億年も早く現れている。そこで、現在のハエとクモの比較はカンブリア紀の節足動物を知るためのひとつの「窓口」となりうると考えたのである。
クモ選択のもうひとつの重要な理由は、クモが胚発生を研究する上で他の生物にはない多くの利点を持っていることである。特に、初期胚の観察が容易である点と多数の卵を計画的に採取できる点は、発生の研究でよく使われているウニやホヤなどと比べてもひけをとらない【図2】。

(図2)新しい実験動物「オオヒメグモ」
人家や山などに普通に見られるクモ。からだは、6~8mm。受精後、母親グモは卵嚢をつくり、そこに卵を産み付ける。
そしてさらに重要な理由は、ハエとクモは比較が可能でありながら、違いが歴然と存在していることである。その違いの本質を追求することで、節足動物の祖先の状態を吟味し、他の動物門との関係性を見出したいと考えている。ではハエとクモはどこが違うか。
3.ハエとクモの違いに注目
私たちが使っているオオヒメグモの卵はみごとな球形である。発生が始まると卵の表層でに細胞が現れ、その細胞の増殖と移動により胚盤と呼ばれる帽子状の細胞のシートが形成される。この時点の胚は放射相称で、将来、胚盤の中心が後(尾部)に、周縁が前(頭部)になることはわかっているが、背腹軸は予見できない。一方、ショウジョウバエの卵はフットボールのように細長く、その形からも遺伝子発現からも将来の前後・背腹を予見できる【図3】。背腹軸については、受精前に準備されたDosalタンパク質がその軸に沿って濃度勾配をつくっており、発生が進むと濃度の高い腹側から中胚葉が形成され、背側に向かって順に、腹側正中細胞、神経組織/腹側表皮、側方表皮、背側表皮、胚外組織に領域化される。この領域化は、Dorsal蛋白質が濃度によって異なる遺伝子の発現を制御することで行われることもわかっている【図4】。
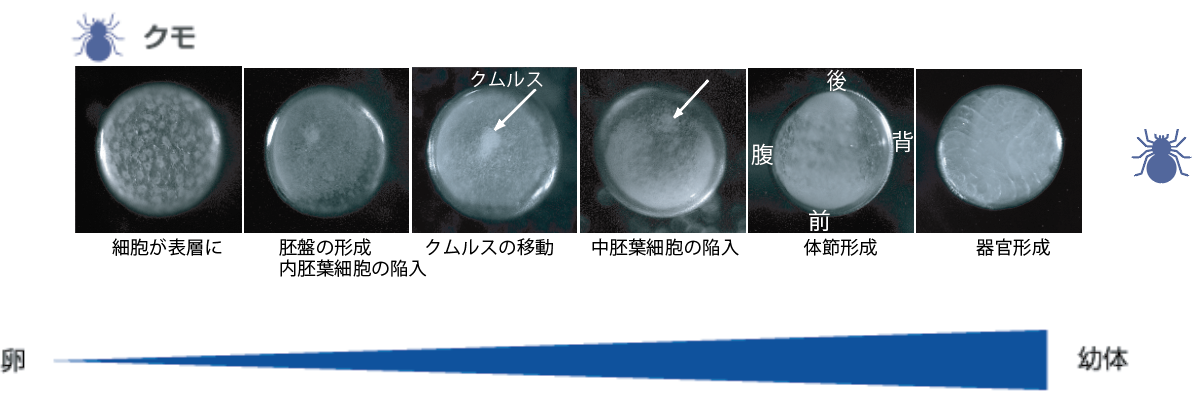

(図3)ハエとクモの胚発生の顕微鏡写真
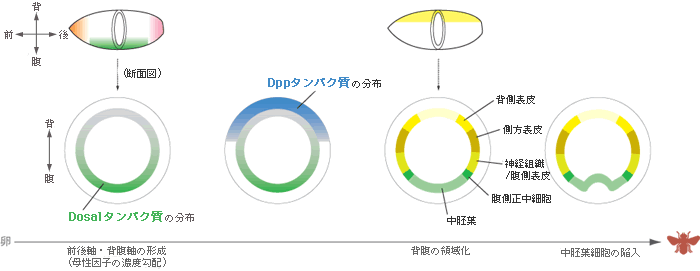
(図4) ショウジョウバエのDorsalタンパク質による領域化
ショショウジョウバエの発生では核だけが先に分裂し、細胞の仕切りは後からできる。この多核性の胞胚の中では、受精前に準備される母性因子が不均等に分布する(■■■)。フットボール型の縦方向に濃度勾配をなすDorsalタンパク質は、断面図では下側の領域で最も濃度が高く、上側にいくにつれ低くなる。発生が進むとDppタンパク質がこのDosalタンパク質の濃度勾配に基づいて発現し、背側領域の形成を誘導する。一方、横方向に濃度勾配をつくるBicoidタンパク質は、体の前後の領域化にはたらく。
ショウジョウバエと同様の背腹軸形成のしくみは甲虫の一種でも働いていることが示されているが、昆虫以外の節足動物では全く報告がなかった。
ではショウジョウバエと違って形の上では放射相称であるクモ胚は、胚盤を形成した後、どのようにして体軸を作るのだろうか? ここからが観察していて面白いところだ。クモ胚を見ていると、胚盤の中心が膨らんで、それが移動し始める。クムルスと呼ばれるこの膨らみの移動した領域が背側になり、引き続いて起こる細胞の再配列によって前後軸、背腹軸が形となって見えるようになる。実は、50年も前にHolmは、別種のクモでクムルスを移植して双子の胚を作り、クムルスが体軸を作る上で重要なはたらきをしていることを示唆していた【参照:図3のクモ胚、クムルスの移動】。
私たちは、ショウジョウバエの体軸形成にはたらく遺伝子と相同な遺伝子をクモから単離して、それらの遺伝子の発現と細胞の挙動を解析した。その結果、胚盤は遺伝子発現から見ても放射相称であり、中心を後方、周縁を前方とする前後軸の位置情報がセットされていることがわかった。一方、背腹軸の形成に重要なクムルスの膨らみが、胚盤の中心に由来する10個ほどの間充織細胞の塊であることも確認した。さらなる解析で、これらクムルス間充織細胞は、シグナル分子Dppを発現しながら胚盤の内面を移動し、上皮細胞に対してDppシグナルを伝達していることが明らかになった【図5】。
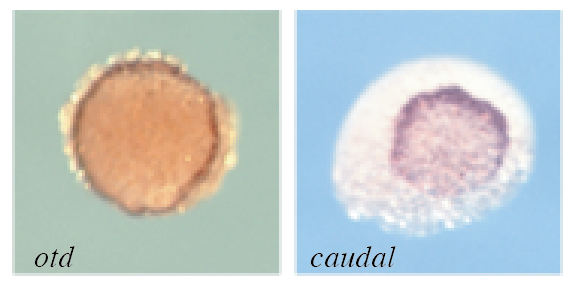
(図5-1) クムルスと遺伝子解析 (1)クモ胚の前後軸形成に関わる遺伝子発現
ハエの前後の領域を決めるotd/caudal遺伝子と相同な遺伝子が、クモの胚の前後の両端に発現。
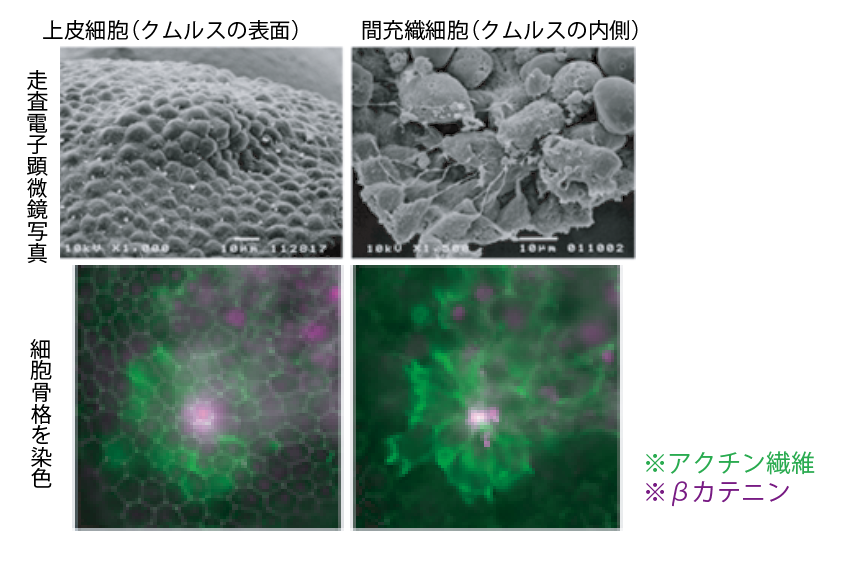
(図5-2) クムルスと遺伝子解析 (2)クムルスにおける細胞の様子
胚盤に出現する上皮細胞の隆起(クムルス)は、その直下に存在する10数個の移動性の間充織細胞によって引き起こされる。
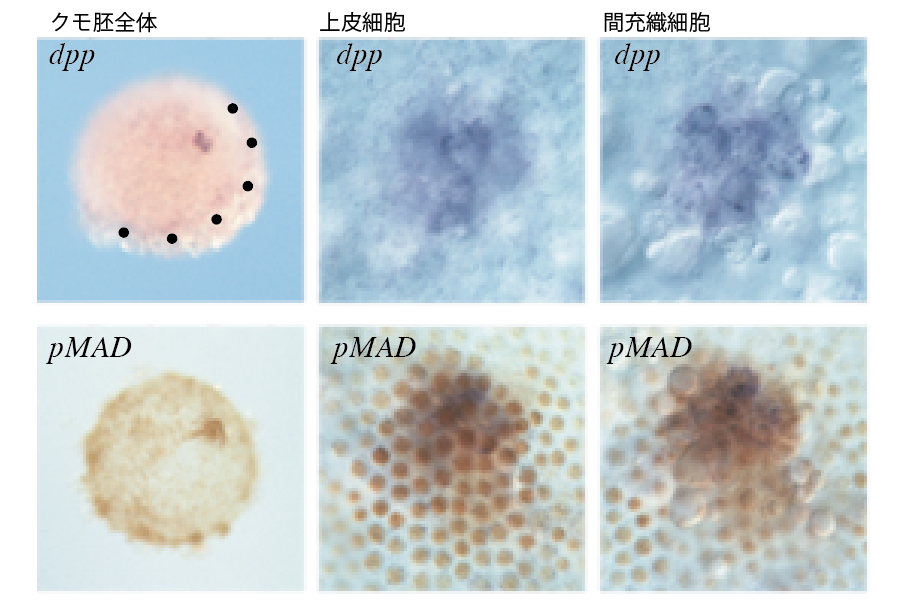
(図5-3) クムルスと遺伝子解析 (3)クムルスでの遺伝子発現(胚盤形成後)
クムルスの内側の間充織細胞がシグナル分子Dppを発現しながら移動。近傍の上皮細胞に背側領域を決める情報を伝えていく。
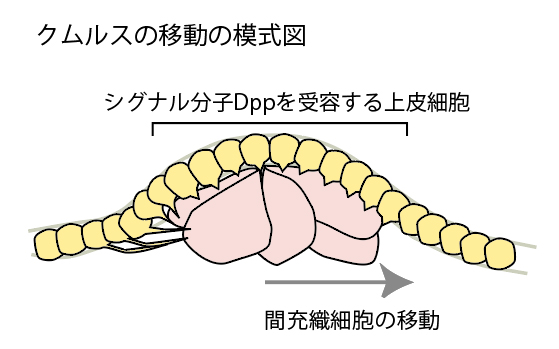
(図5-4) クムルスと遺伝子解析 (4)クムルスの移動の模式図
Dppシグナル分子はハエ胚で背側領域を誘導していることがわかっているので、クモ胚でもDppが同じ役割を果たしていると考えると非常によく辻褄が合う。それではどこにハエとクモの違いがあるのだろう? ハエ胚では予めセットされたDorsal蛋白質の濃度勾配が背腹軸を作ることは前に述べたが、Dppはその濃度勾配に従って背側領域に発現する。それに対してクモ胚では、Dppの発現する領域は予め決まっておらず、Dppを発現する細胞(クムルス間充織細胞)が発生初期に出現し、移動することによってはじめて、背腹軸を作るのである。この細胞移動の方向を決定する仕組みはまだ分かっていないが、ハエとは異なる背腹軸形成のしくみが存在することは明らかである【図6】。
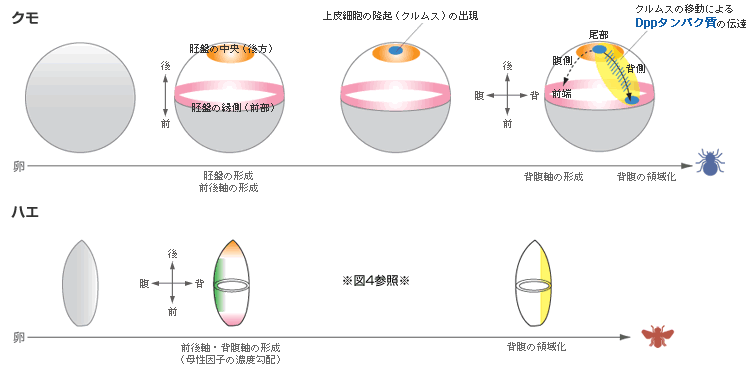
(図6)ショウジョウバエとクモの軸形成の相違
ハエとクモの共通点は、節足動物の祖先においてもDppシグナル分子が胚の背側領域の決定に使われ、この状態が子孫に引き継がれたことを意味する。一方両者の相違点は、祖先からハエやクモに向かった進化において、背腹軸を形成する発生プログラムのどの部分に変型(バリエーション)を持ち得たのかを教えてくれる。
4.まだ始まったばかり
クモの研究はまだ始まったばかりで、左右相称動物の進化のメカニズムを知りたいという私たちの思いにはまだまだ手が届かない。現在、クモの中胚葉がどこからどのように発生するのか、クモの外胚葉の腹側正中線がどのように決定されるのか、クモ胚の後方の尾葉と呼ばれる領域でどのように体節が形成されるのかを調べている。これまでの結果でクモがハエと脊椎動物を橋渡しする貴重な存在であるという手応えを感じている。
動物の進化の復元をどこまで論理的に進められるのか、とりあずクモを使って試してみたい。
さらに詳しく知りたい方へ
- Akiyama-Oda, Y. and Oda, H. (2003). Early patterning of the spider embryo: A cluster of mesenchymal cells at the cumulus produces Dpp signals received by germ disc epithelial cells. Development 130, 1735-1747.
- シリーズ進化学4「発生と進化」
編集:佐藤矩行、野地澄晴、倉谷滋、長谷部光泰(岩波書店)
秋山-小田康子(あきやま-おだ・やすこ)
科学技術振興機構 さきがけ研究21 研究員
1998年東京大学大学院理学系研究科博士課程修了。日本学術振興会特別研究員などを経て、2001年より現職。同年よりJT生命誌研究館、小田ラボにて研究。
小田広樹(おだ・ひろき)
JT生命誌研究館 研究員
1996年京都大学大学院理学研究科博士課程修了。科学技術振興事業団月田細胞軸プロジェクトグループリーダーなどを経て、2001年より現職。














