RESEARCH
「語る科学」
メダカ-日本が育てたモデル動物
今メダカが注目されている。ゲノムプロジェクトが始まり、脊椎動物のかたちづくりを調べるモデルとして変異体の解析も進行中だ。研究室で飼育されているメダカも、元をたどれば日本の各地域で生息していた野生メダカの子孫たち。長い歴史を持つ日本のメダカ研究は、科学の中にも、日本の文化、日本人を感じることがあることを気づかせてくれる。
1.メダカ --- 古くて新しい実験動物
動物のからだがどのように作られるかを調べるには、まずは研究対象の生きものを決めなくてはならない。サカナはタマゴから生まれるので、最初の受精卵の様子から観察するのに適している。実験室で飼うことを考えると、卵は観察しやすい大きさがいいが、体はなるべく小さなほうがよい。サカナは親の大きさと卵の大きさにあまり関係がなく、例えばマグロの卵はサケの卵(イクラ)よりも小さい。そこで、小型魚類の出番となる。
実験動物としての小型魚類の代表は、ゼブラフィッシュとメダカだ(図1)。これらの特長は、まず飼育が容易で毎週100~200個の卵を産み、しかも2ヶ月ほどで成長する短い世代サイクルなので、遺伝学研究に向いている。さらに、胚が透明で発生速度が速く、実験発生学の対象としても優れている(図2)。また基本的に哺乳類と同等の器官・組織を備えており、今ではいくつかの変異体がヒトの遺伝子疾患のモデルとして研究され始めている。

(図1) メダカとゼブラフィッシュ
写っているメダカ(上)は、体色の変異体の一つであるヒメダカ。
ちなみにゼブラフィッシュは、昔からゼブラダニオという名で売られている初心者向けの熱帯魚である。
ゼブラフィッシュの研究者だった私がメダカに興味を持ち始めたのは5年前。当時ゼブラフィッシュの研究者人口はうなぎのぼり、大きなプロジェクトが欧米で立ち上がり次々と新しい成果が専門誌に発表されていた。その研究基盤の多くが欧米で作られ管理されている状況になんとなく不満と不安を持ち、メダカのこれまでの研究を調べてみるとメダカの利点が次々と見えてきた。
メダカは東南アジア原産で、日本の水田ではどこでも群れており、江戸時代にはすでにペットとして庶民の家庭に入り込んでいた。当然のことながらメダカ研究の大部分は日本産である。特筆すべきことのひとつは、先輩研究者たちの努力によって交配可能な北日本集団由来と南日本集団由来の純系(註1)が樹立されており、しかもこれら南北系統間のゲノムを比較すると遺伝子のコード領域で約1%、非コード領域で約3%の塩基が異なっている(図3)。これはゼブラフィッシュの異なる系統間でみられる違いの5倍程度で、ヒトとチンパンジーという種を越えたゲノムどうしの比較でもその差は1%強と考えると、メダカの種内変異の幅は極めて大きい。高頻度の多型を示す純系の存在は、連鎖地図作成にもちいる多型マーカーの単離や、突然変異体の原因遺伝子の位置を染色体上で探す場合に特に有利である(註2)。一言で言えば遺伝学研究がやりやすいということだ。さらに、ゲノムサイズが比較的小さいことや生育温度が幅広いことなど、実験動物としてゼブラフィッシュと同等以上の可能性を持つと確信し、メダカ研究を本格的に始める決心をした。
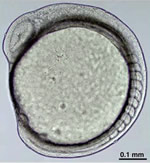
(図2) 透明な小型魚類の胚
初めて透明な受精卵を見たときのゾッとするような衝撃、本当に美しいと思った。
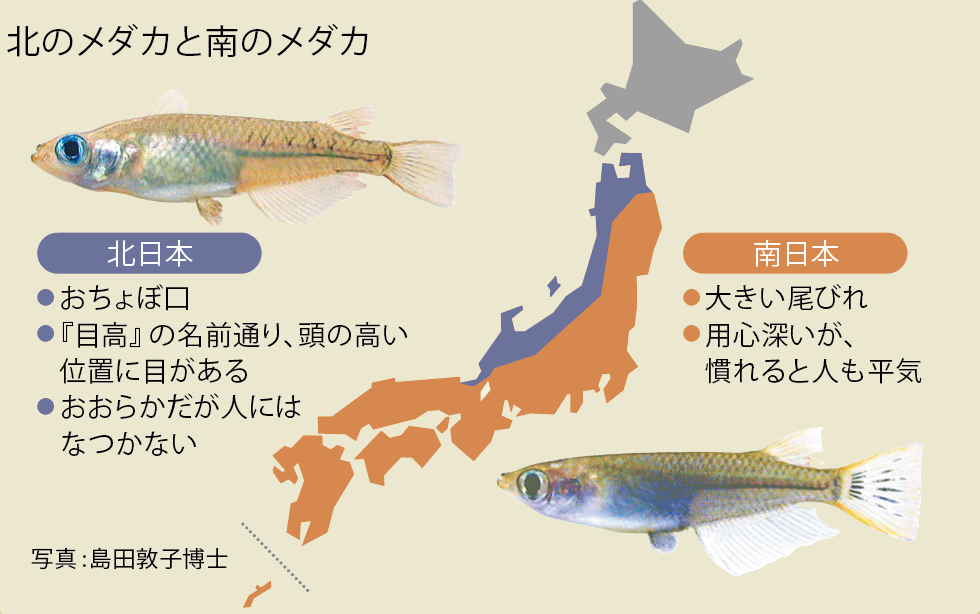
(図3)北のメダカと南のメダカ
南北集団間では、顔つきや体型、性格も異なる。この個性の違いがどんな遺伝子の影響下にあるのかの探索も始まった。
(註1) 純系
兄妹交配をくりかえすことで個体間の遺伝的なばらつきがほぼなくなった系統。近交系とも呼ばれる。野生集団から純系を作ろうとする場合、ヘテロに持っていた劣性致死遺伝子の影響が現れるなど困難な場合が多い。
(註2) 多型マーカー
ゲノムDNAの同じ部位にある塩基配列が個体間で違うことを多型が存在するといい、異なる配列部分を多型マーカーと呼ぶ。ヒトの血液型にA,B,AB,Oと個人差があるのも、血液型を決める遺伝子に多型が存在するためである。多型には、遺伝子の機能に影響を及ぼすものも及ぼさないものもあるが、どちらもメンデルの法則にしたがって遺伝する。純系個体では両親から受け継いだ2組のゲノムDNAに塩基配列の違いはないが、異なる系統(例えば北集団のオスと南集団のメス)の純系を掛け合わせると、 一世代目は北集団の染色体と南集団の染色体を1組ずつ持った個体が生まれる。この個体の生殖細胞が減数分裂を行うと染色体の組換えがおこるので、一つの染色体に北由来、南由来の部分が混ざることになる。染色体のどこまでが北由来でどこまでが南由来かは、それぞれの系統が持つ多型マーカーが出現するかどうかを調べればよい。染色体のどこで組換えが起こるかは偶然で決まるので、多数の組換え体(すなわち二世代目の個体)を解析すれば、染色体上にどのように多型マーカーが配置しているかを知ることができ、このようにしてでき上がった染色体の地図を連鎖地図と呼ぶ。同様のことを、例えば南系統に由来する突然変異体と北系統の野生型の掛け合わせで行えば、変異したDNA配列がどの多型マーカーの近くにあるかがわかり、原因遺伝子を突き止めることができる。
2.メダカの変異体を単離する --- かたちづくりの理解をめざして
ある遺伝子を不活性化したら、表現型がどう変わるかを見て遺伝子の機能を探る逆遺伝学が現在の研究の主流だ。面白い表現型が現れれば機能解析が順調に進むが、あらかじめ対象とする遺伝子が判明していなければならない。現時点では、ゲノムの全配列が判明したとしても、数万種類にのぼる遺伝子産物をすべてこの手法で解析することは現実的ではない。従って、まず注目する現象に的を絞り、その現象に変化が現れる変異体を探す古典的な(順)遺伝学が、生命現象の素過程を明らかにする上では依然として有力な手段だ。私たちは脊椎動物のからだづくり(個体の発生)を支配する重要な遺伝子群を明らかにする目的で、2000年から国立遺伝学研究所で、2002年からは一部東京大学、東京工業大学(工藤明研究室)とも共同で、発生に異常を示すメダカ変異体の単離(スクリーニング)を進めている。このような計画はもちろんゼブラフィッシュで既に試みられているが、メダカとゼブラフィッシュでは遺伝子の構成がかなり異なっているらしく(後述)、ゼブラフィッシュでは見られない表現型を持つ変異体が多数単離できた。
現在進めているスクリーニングは、外形・内臓器官など形態異常が簡単にみつかるものが中心だ。例えば、胴体部が変形する変異体の中に脊椎骨が異常な変異体がある。極端な例がtoguro(tgr)だ(図4)。骨を染めてみると一目瞭然で、tgr変異胚では骨がまったく形成されていないことがわかる。さらに興味深いことに、tgrは温度感受性(註3)を示しており、30度以上の高温飼育下で異常が強く、24度以下の低温ではほぼ正常に近い。重要な遺伝子は発生、成長の様々な段階で使われていることが多く、単なる機能欠損では発生初期に現れる影響だけが目立ち(ここで死んでしまう場合も多い)、それ以降の異常がどこまで変異遺伝子の直接の結果か判断しにくい。生育温度の変化だけで表現型の出現時期が制御できる温度感受性変異体の単離はショウジョウバエ、線虫などでは有力な手法となっており、脊椎動物の発生研究でもその応用が期待されていた。しかし恒温動物である哺乳類ではそもそも不可能な試みであったし、熱帯性のゼブラフィッシュでも生育許容範囲が22―30℃と狭いので温度変化の再現性が難しかった。生育温度が4―40℃と広い温帯性のメダカならではの成果である。

(図4)とぐろを巻くメダカ
正常なメダカの稚魚(上)と変異体「とぐろ(toguro)」(下)
「とぐろ」では背骨などの骨(赤く染まっている部分)が全く形成されていない。この変異は温度感受性を示し、生育温度の変化で異常の起きる時期を制御できる。
また、メダカ胚はゼブラフィッシュ胚よりもさらに透明度が高く、外から内臓器官が良く見える。注目している内臓器官の変異体には、肝臓・甲状腺の発生異常、内臓逆位(左右軸の異常)などがあり、ヒトの病気を連想させるものも多い(図5、図6)。脊椎動物の発生機構の共通性(保存性)を考えると、これらの変異体の研究からヒトを含めた脊椎動物の重要な発生機構や疾病の原因が明らかになると期待している。次に計画中のスクリーニングでは、胚の透明さをさらに有効に利用するために、内臓器官の一部を蛍光タンパク質で可視化できるトランスジェニックメダカ(註4)を作った(図7)。このメダカを用いれば、さらに多くの内臓器官の変異体を単離できるはずだ。

(図5)内臓の発生異常
正常個体(上)と変異体(下)。赤線で囲った部分が正常な肝臓。変異体では、肝臓の細胞が変性を起こしたため(赤矢印)、黒く見えている。
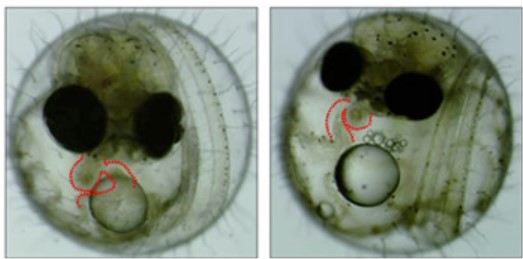
(図6) 内臓逆位の変異体
心臓(赤色の破線で囲った部分)のかたちが異常になった変異体(右)。正常個体(左)とは心臓の折れ曲がる方向が逆になっている。
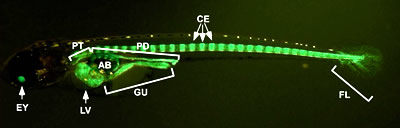
(図7)内臓器官が光るトランスジェニックメダカ
紫外線を当てると体の特定の場所が光るように操作したメダカ。この個体では、肝臓(LV)、消化管(GU)、腎管(PT、PD)、椎骨(CE)が主に発光している。
3.魚の体型を制御する遺伝子、Double anal fin (Da)
ここで、魚類の形態進化を考える上で興味深い変異体を紹介する。野生集団から研究者が単離した変異体の中にDouble anal fin (Da)という変異体がある(名古屋大学分子応答センターで収集)(図8)。その名の通り、背びれが尻ビレ(anal fin)の形に変わっているので、2つの尻ビレを持っているように見える。通常メダカは池や川の表層を泳ぎ、落ちてきた昆虫などを餌としている。その生活スタイルに合うように、メダカの体型は背側が平坦で体色は黒く、逆に腹側は銀色である。Da変異体は体全体がtear-drop型(涙型)、これはマグロやカツオのように海洋の中層を回遊する魚の特徴だ。また背側にも腹側同様に銀色の色素が分布しており、泳いでいる姿を上から見るときらきら光を反射してよく目立つ(自然界では見つかりやすくとても生きられないが・・・)。つまりDa変異体は、頭部を除く体全体の色やかたちが腹側鏡像対称となっているのだ。ちなみに近ごろペットショップで「ひかりメダカ」という名前で売られているものは、どうもどこかの研究室から流出したDa変異体らしい。魚類では、近縁種であってもその生息環境(表層、中層、底層)により体型が大きく異なっている場合が多く、このDa変異体の原因遺伝子はそのような体型を司る遺伝子ではないかと考えられている。

(図8)体型の変異体Da(通称“ひかりメダカ”)
普通のメダカの胴体は、背ビレのある平たい背中(橙色)と尻ビレのある腹側(緑色)でできている(上)。たった一つの遺伝子の変化で、背中が腹側の構造になり、マグロのような体型のメダカになった(下)。
最近、東海大学の大塚正人博士の長年の努力により、この原因遺伝子がZicと呼ばれる遺伝子であることが分かった。幸いこの遺伝子の機能解析を手伝う機会があり、大変楽しませてもらった。Zic遺伝子はマウスで最初に報告されたもので、神経系、体節(筋肉、骨の原基)など広い範囲で胚発生期に発現している。ただし、Zic遺伝子を働かないようにしたマウスでは、腹側鏡像対称にはなっていない。Da変異体では、遺伝子から作られるタンパク質そのものには変化がなく、体節の背側での発現だけが消失していることが判明した。このため背側の筋肉が減少し、その影響が表皮にまで及んでヒレの形から色素の分布パターンまで変化したらしい。Da変異体の存在は、メダカの祖先の魚の基本型が中層回遊型のtear-drop型(涙型)であることを想像させる。この例は、最近よく言われる、「進化はタンパク質をコードしている領域(この変異は致死になり易い)よりは制御領域の変異の積み重ねでおこる」という考えを見事に支持しているように思われる。
(註3)温度感受性変異
突然変異体の中には、ある条件下でだれ変異表現型を発現するものがあり、このような変異を条件変異とよぶ。温度感受性変異では、変異体がある温度以上の高温(もしくはある温度以下の低温)にさらされたときにだけ、表現型を示す。
(註4) トランスジェニックメダカ
「トランスジェニック」とは、遺伝子DNAを人工的な方法で細胞の中に入れ込むこと。これにより受精卵のゲノムに外来DNAを組み込ませることができれば、成長した個体の全ての細胞が外来DNAを持ち、生殖細胞を通じて次世代にも伝えられる。本文で紹介されたトランスジェニックメダカは、「紫外線を当てると内臓が緑色に光る」という性質の遺伝子をメダカゲノムに組み込み、簡単に内臓器官の発生が観察できるようにしたものである。
4. メダカゲノムプロジェクト --- 魚類進化の道筋をたどる
ヒト、マウス、ラットなど哺乳類のゲノム配列の決定に続き、小型魚類でもゲノムプロジェクト(ゲノムDNAの全塩基配列の決定)が進行している。魚類の中で先行していたのが、トラフグ、ミドリフグ、ゼブラフィッシュの3種である。ゲノムサイズの小さいフグは概要配列が既に報告されているが、実験室内での大量飼育が難しく発生学・遺伝学への応用がきかない。ゼブラフィッシュはゲノム構造の複雑さに加えて純系を使用していないためにプロジェクトが難航しているらしい。ここでメダカの出番となる。
メダカは多様な硬骨魚類の中で、系統的にゼブラフィッシュとフグの中間に位置し、ゲノム進化を考える上で重要な存在だ(図9)。ゼブラフィッシュとフグ・メダカの祖先は約2億年前に分かれ、メダカとフグは約6千万年前に分岐した。ゲノムサイズも8億塩基対と、ゼブラフィッシュ(17億塩基対)とフグ(4億塩基対)のほぼ中間である(ちなみにヒトは30億塩基対)。魚類は進化の初期にゲノム重複を起こし、主要な遺伝子の数は哺乳類の2倍近くになった。その後、進化的に新しく登場した魚ほどゲノムサイズを減らしていったと思われる。ゲノム重複を起こした生物の遺伝子には、余分な遺伝子の不活性化や役割分担、新しい機能の獲得などが頻繁に起こると言われている。系統の離れたゼブラフィッシュとメダカでは、遺伝子の構成がかなり違っていると予想される。
我々は、国立遺伝学研究所・小原雄治研究室、東京大学・森下真一研究室と共同でメダカゲノムプロジェクトを計画し、2002年秋から配列決定を始めた。用いたのは、終戦直後に愛知県弥富町で売られていたヒメダカとシロメダカの子孫に由来する、Hd-Rrと呼ばれる南日本系統の純系メダカ。2004年末までに概要配列の完成を目指しており、変異体の原因遺伝子の単離が飛躍的に加速するだろう。近い将来、多様な硬骨魚類の進化が刻まれているゼブラフィッシュ、メダカ、フグのゲノム配列と構造の情報が手に入る。ゲノム配列が判明しているヒト、マウス、ラットとの詳細な比較が可能になり、ゲノム進化、種分化研究の新たな地平が見えてくることは間違いない。
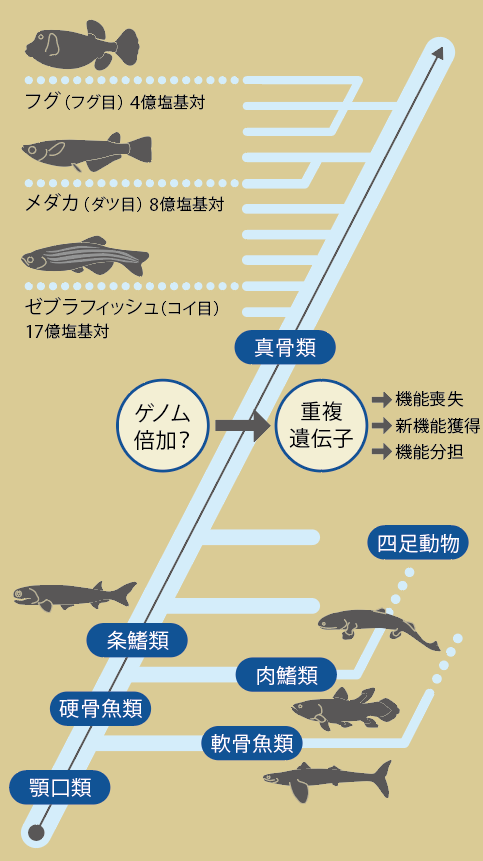
(図9)魚類の系統とゲノムサイズ
現生魚類の大半を占める真骨類は、進化の初期にゲノム重複を起こしたらしい。その後、重複した遺伝子の欠失や役割分担が起こった。
5.メダカの棲んでいる環境
私が子供の頃は、メダカは日本の水田や用水路にあふれていた。まさか絶滅危惧種に指定される(1999年環境庁レッドリスト)とは誰も想像だにしていなかっただろう。日本のメダカ研究が、各地域に生息するメダカの多型や種分化を対象とするものが多かったのは、メダカがどこにでもいる反面、各水系に依存した分化傾向を持っていたためと思われる。読者には是非、日本を含む東南アジアのメダカ族の系統と地域集団、種分化についての膨大な研究も参照していただきたい(註5)。
メダカを研究するようになってから、私の興味は研究室の枠を超えて拡がったような気がする。研究室の中で飼育されているメダカも元をたどれば日本の各地域で生息していたメダカの子孫たちである。多くのメダカ系統は最初に採集された日時と場所まで遡れる。目の前の水槽で泳いでいるメダカと絶滅の危機に瀕する水田のメダカが私の中で重なってくる。メダカが日本の自然と人間の関係の歴史を教えてくれるからかもしれない。科学の中にも、日本の文化、日本人を感じることがあることに気づいた。日本の自然を生かした研究で世界に通じる面白い成果をあげてみたい。
(註5)さらに詳しく知りたい方へ
● すべての情報はこのサイトから、MEDAKAFISH HOMEPAGE
http://biol1.bio.nagoya-u.ac.jp:8000/
● 『メダカと日本人』
岩松 鷹司 (著)
出版社: 青弓社 ; ISBN: 4787232010 ; (2002/05)
● 『メダカ学』
岩松 鷹司 (著)出版社: サイエンティスト社 ; ISBN: 4914903105 ; (1993/09)
武田洋幸(たけだ・ひろゆき)
1958年生まれ。1985年東京大学大学院理学系研究科博士課程退学。理学博士。東京大学理学部助手、理化学研究所研究員、名古屋大学理学部助教授、国立遺伝学研究所教授を経て現在東京大学大学院理学系研究科教授。