Experiment
遺伝子の中の厄介者,イントロンはどうしてなくならないか
私たちの遺伝子のほとんどにあるイントロン。遺伝子が動く際には切り捨てられ使われない不思議な存在です。一見,不要に見えるイントロンはなぜなくならないのでしょう。JT生命誌研究館での研究から生まれた,大胆な仮説を紹介します。
イントロンの不思議
生物を見ていると,びっくりするほど巧妙な仕組みがある一方で,どうしてこんな無駄があるのかと思うような不思議な現象に出会う。
ヒトゲノムは30億塩基対のヌクレオチドが並ぶといわれるが,その中でタンパク質を作る情報をもつ遺伝子の部分は数%しかない。そのうえ,遺伝子の中にも,タンパク質を作るための情報をもたず,RNAに転写された後に切り捨てられる部分がある。それがイントロンだ(図①)。必要な情報はエキソンと呼ばれる領域に断片的に存在しており,それがRNAの段階でつなぎ合わされ,さらに化学的に修飾を受けて,タンパク質合成(翻訳反応)の鋳型となるmRNAができる。
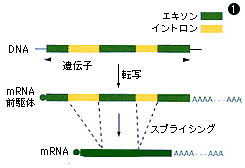
イントロンとエキソン
①遺伝子の中にあって,タンパク質を作るための情報をもつ部分がエキソンで,もたない部分がイントロン。イントロンは,遺伝子が働く際に切り取られ捨てられる。
私が初めてこの話を知ったのは,生物学科の学生の頃で,今から20年も前のことだ。その頃研究に使っていたバクテリア(原核生物)にはイントロンがない(当時はそう思われていた)のに,ヒトを含む真核生物ではなぜこんな手間暇のかかることをするのかとても気になった。さらに,当時話題になっていた逆転写酵素の働きで,mRNAがDNAに変換(逆転写)され,ゲノムに埋め込まれて生じたと思われるイントロンをもたない遺伝子(processed gene)が,細胞の中で立派に機能していることも知られていた。イントロンがなくても遺伝子は働けるのに,なぜイントロンはゲノムから根絶されないのだろうか。20年たってやっと,思いもよらないところからこの問題に取り組み,解答を得ることになった。
イントロンには役割がある?
JT生命誌研究館で私たちのグループが研究を始めるまでに,すでにイントロンについては多くのことがわかっていた。たとえば,DNAから転写されたばかりのRNAの配列を正しく認識してイントロンの部分を除く(スプライシング反応という)には,RNAとタンパクの複合体からなる大きな専用の装置「スプライソゾーム」が必要であることが明らかにされた。また,進化の過程で複数のエキソンの新しい組み合わせが登場し,それまでにない酵素ができる場合,イントロンがノリの役割をして新しい組み合わせがうまく機能する率を高めていることもわかった。そのほか,イントロンが遺伝子の転写活性を強める例や,イントロンの切り出し部位を変えて遺伝子の発現調整をしたり,異なる酵素活性をもつタンパク質を作る例も知られている。
こうして,少なくとも一部のイントロンは何らかの機能をもつことがわかったのだが,これで長年のモヤモヤもすっきり,やはり生物に無駄はないと思ったのでは,イントロンの本質を見誤るというのが,われわれの研究である。
核型イントロンの起源
- オルガネラにあるgroupIIイントロン
まず,イントロンが真核生物に特有のもので,細菌(原核生物)には存在しないという教科書的記述は誤りである(図②)。真正細菌にも,その変形であるミトコンドリアや葉緑体などのオルガネラにもイントロンはある。ただそれは,真核細胞の核ゲノムにあるものとは異なる。
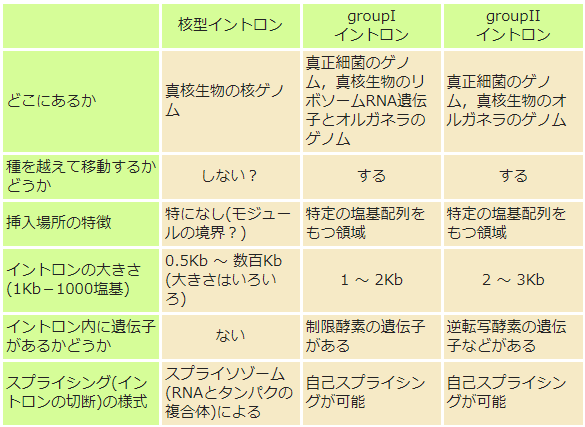
いろいろなイントロン
②真正細菌や,それが共生してできたオルガネラ(ミトコンドリアや葉緑体)にも,核ゲノムにあるものとタイプは異なるがイントロンがある。
それらのイントロンは,groupI, groupIIの2つに分類され,そのうちのgroupIIと呼ばれるイントロンが,謎を解く鍵というのがここでの主題だ。すなわち,この型のイントロンが,真核細胞の核の遺伝子にあるイントロン(核型イントロン)の原型になったという考え(図③)を支持するデータが,蓄積されつつあるのだ。まず,オルガネラのgroupIIと核型イントロンでは,その切り出しやエキソンの結合で見られる有機化学反応がまったく同じである(図④)。さらに,核型イントロンの切り出し装置(スプライソゾーム)に含まれる低分子RNAが,groupIIイントロンの6つの腕に相当するらしい。
そこで,核型イントロンが,細菌やオルガネラにあるgroupIIイントロンの変型であると考えると,生物界における核型イントロンの分布をみごとに説明できる。たとえば,オルガネラをもたない原始的といわれる真核細胞には核型イントロンがない。これらのことから,原始真核細胞に共生した真正細菌がもっていたgroupIIイントロンが,核に入り込んで変型したのが今の核型イントロンであると考えることができる。つまり,原始真核細胞は,オルガネラを得る代わりにgroupIIイントロンに感染し,余分なものを抱え込んだのだ。人生良いことばかりはないとは,生物進化上でも正しい教訓らしい。
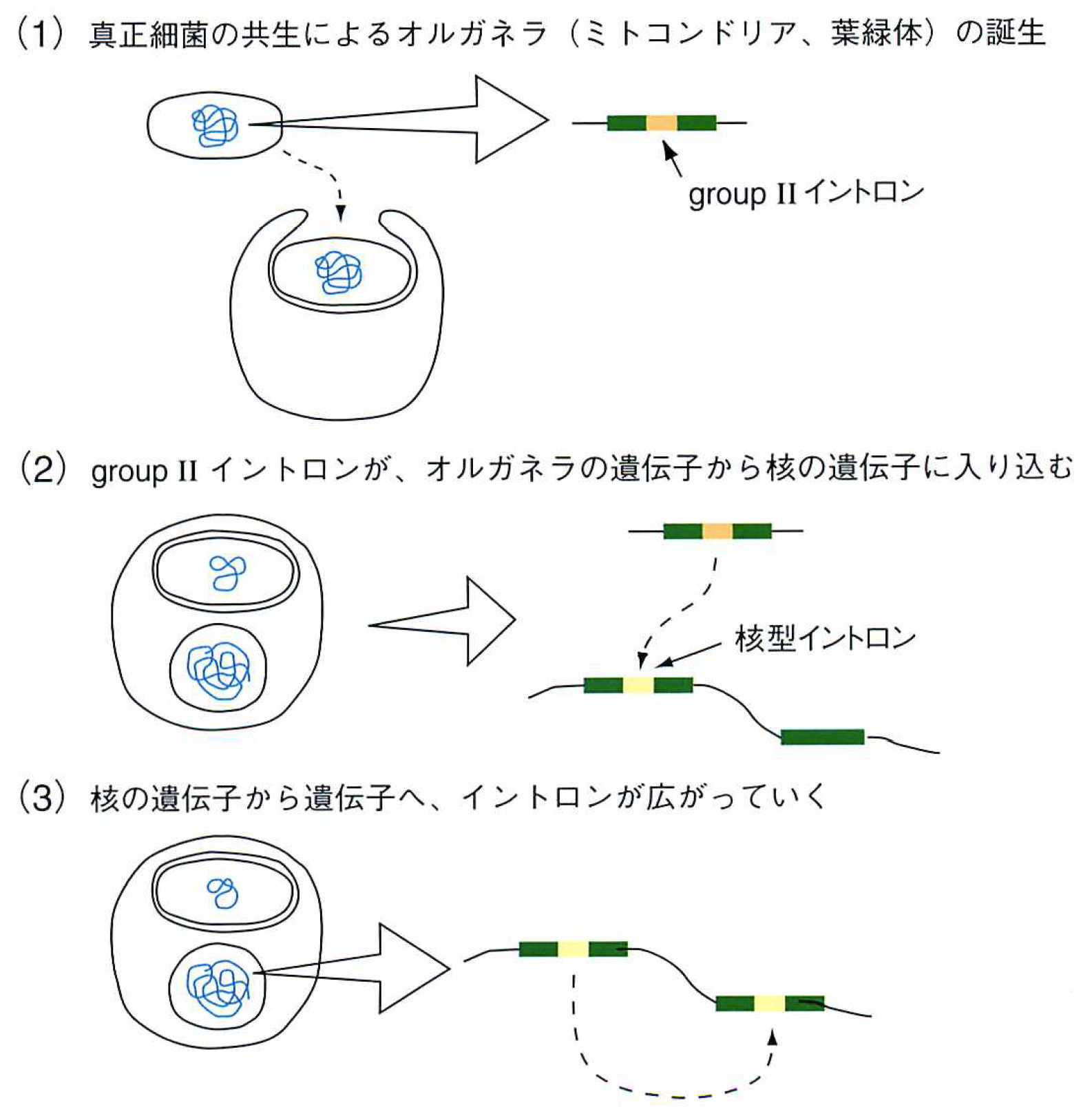
③真核生物の核ゲノム中の遺伝子にあるイントロンは,オルガネラの誕生によってバクテリアからやってきたと考えると,多くの実験データがうまく説明できる。
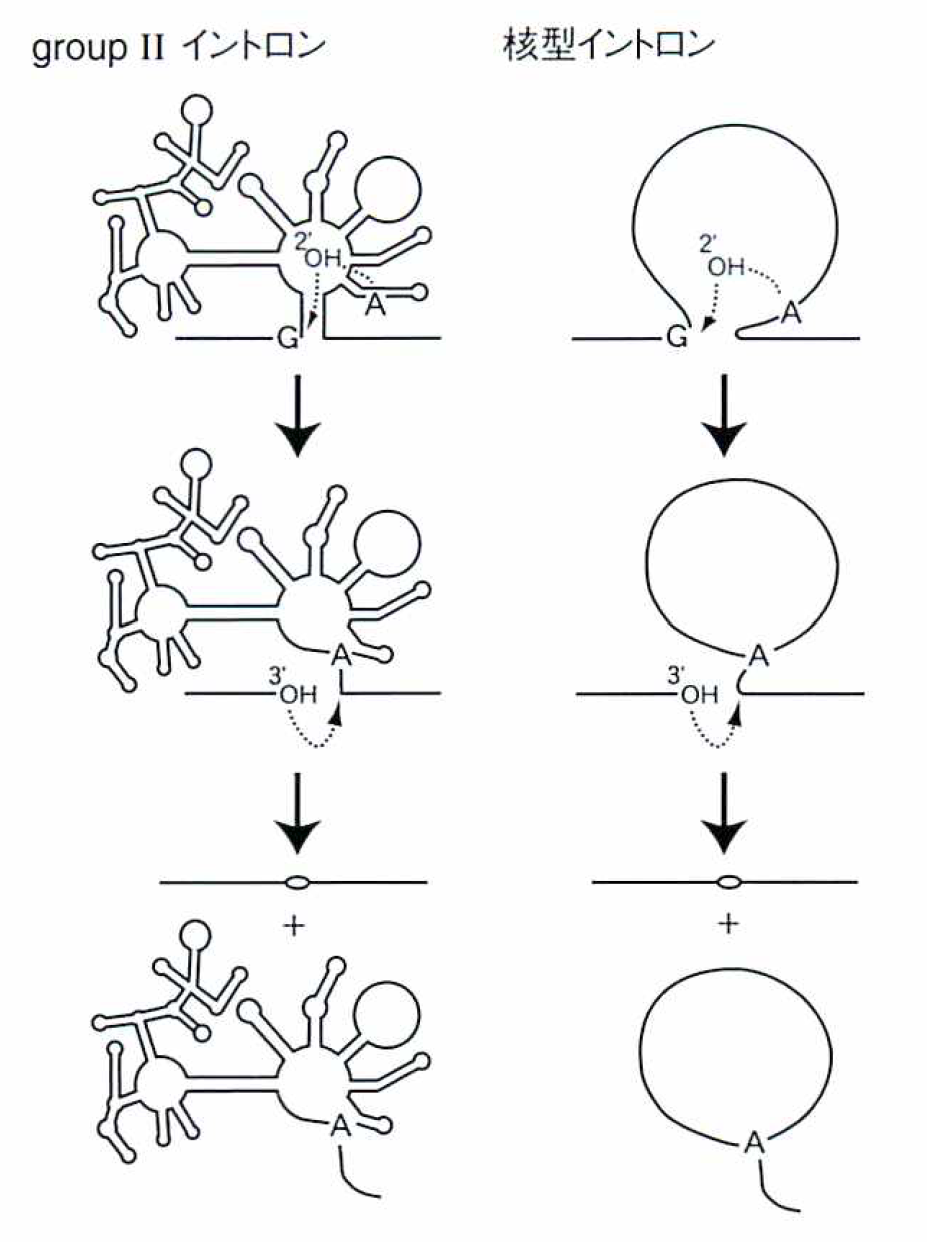
④groupIIイントロンと核型イントロンの切り出し(スプライシング)反応は,有機化学的にはまったく同一である。
動き回る遺伝因子groupIIイントロン
では,核型イントロンの原型ともいえるgroupIIイントロンとはどんなものなのだろうか。groupIIイントロンは,特定の遺伝子に自己のコピーを組み込む能力をもっており,ここに多くの研究者が注目している。オルガネラのgroupIIイントロンは,強い感染性・増殖性を示す,いわゆる利己的な遺伝因子の一つなのだ。
私の研究室の大学院生だった江原が中心になって,ハプト藻のミトコンドリアCox1遺伝子から見いだしたgroupIIイントロンを図⑨⑩に示す。核型のイントロンとは異なり,特異な立体構造をとり,内部には逆転写酵素やDNAを切断する酵素(DNase)をコードする領域がある。こうした酵素の活性を使って,groupIIイントロンはイントロンをもたない遺伝子に部位特異的に自己のコピーを挿入する。
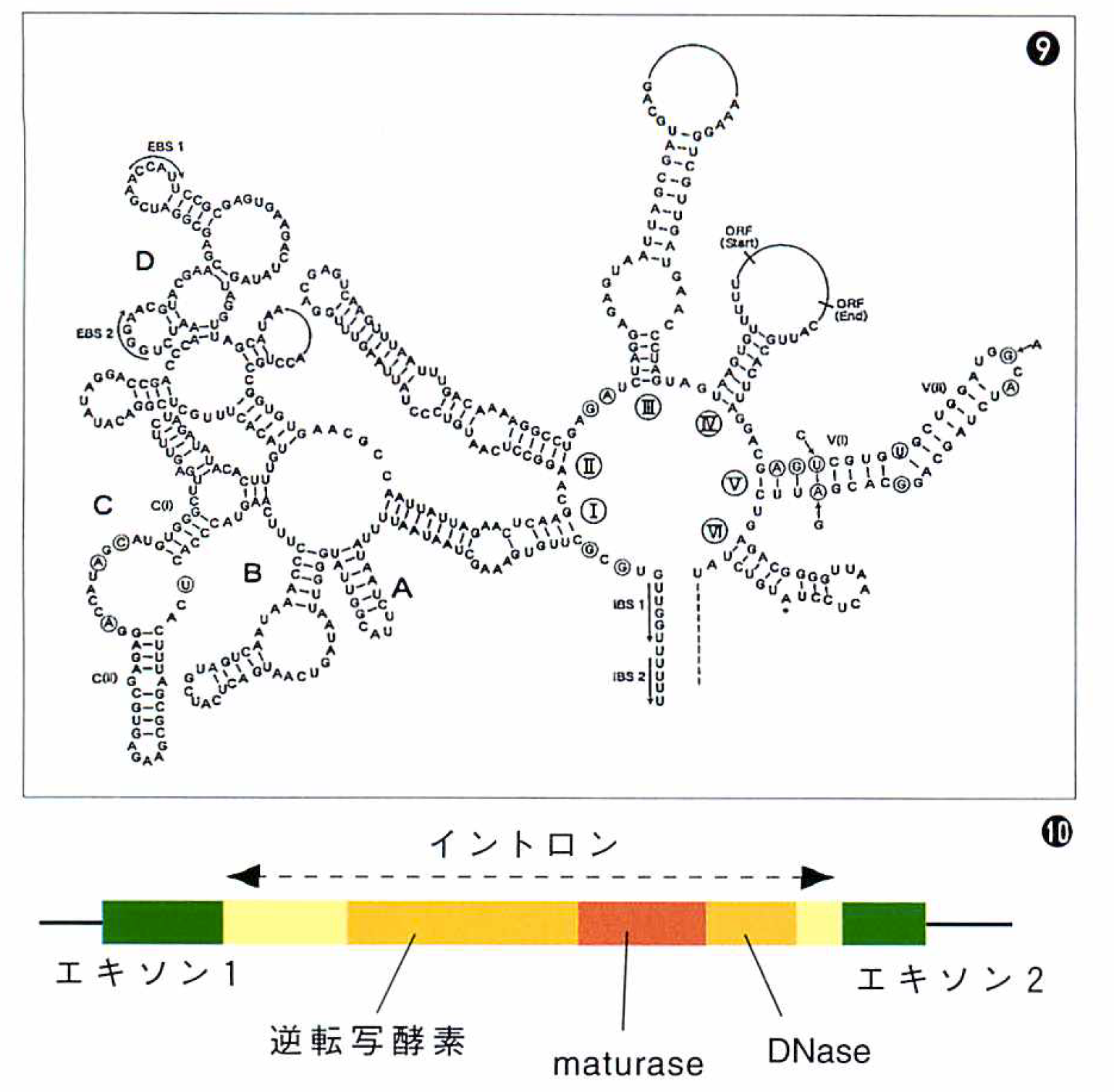
⑨2次構造を示した模式図。右上のループの部分に約1900塩基分のコーティング領域があり,逆転写酵素,maturase, DNaseという3種の酵素の遺伝子がある(3種の遺伝子の位置を示した模式図が⑩)。
機能はまったく不明だが,groupIIイントロンは,ごく稀にではあるが種を越えて異種のオルガネラのゲノムにも転移できるらしい。というのも,イントロン内部の逆転写酵素領域を使って分子系統樹を作成すると,まったく進化系統関係を反映しない樹ができあがるからだ(図⑪)。しかも,同種であれば世界中どの海域から採取された個体も同じイントロンをもつので,集団内(種内)の一匹が特定のgroupIIイントロンに感染すれば,接合で瞬く間に種全体に蔓延するようだ。一方,近縁の種間では同じイントロンをもつ例は見当たらないので,種内に蔓延したイントロンはそれほど長続きせずに失われることがわかる。groupIIイントロンの感染と脱感染は種分化よりも早いサイクルで起こるといえる。groupIIイントロンは,種の壁を越えて動く遺伝因子の一つと考えてよかろう。
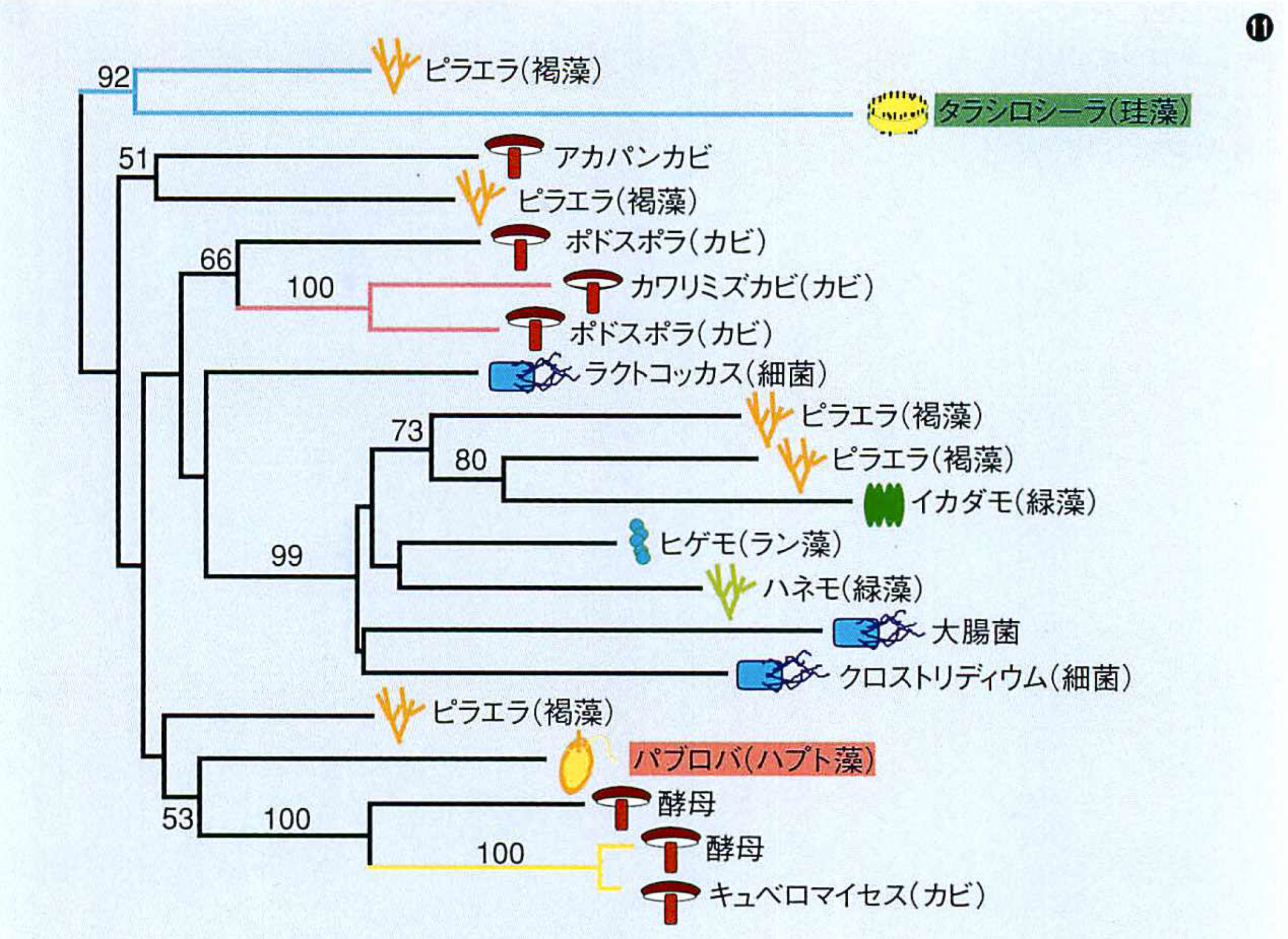
groupIIイントロンは,種を越えて移動する?
⑪イントロンの内部にある遺伝子の配列をもとに作られた系統樹。これを見ると,系統的に遠い生物同士がよく似たイントロンをもつことがわかる。イントロンが種を越えて移動したと考えると説明がつく。
核型イントロンが核の遺伝子に侵入する証拠
こうしたgroupIIイントロンの性質を理解したうえで,核型イントロンの歴史を考えると,次のようなストーリーができる。すなわち,初期の真核細胞の進化途上で,核ゲノムに転移したgroupIIイントロンは,切り出し活性に必須なRNA部や内在酵素を自分自身から切り離して,スプライソゾームという別の装置に仕立てあげた。もとのgroupIIは特定部位にしか侵入できなかったが,核型イントロンとスプライソゾームの組み合わせなら,ゲノムのどこにでも自由に侵入できるので,一気にゲノム中に広がったに違いない。
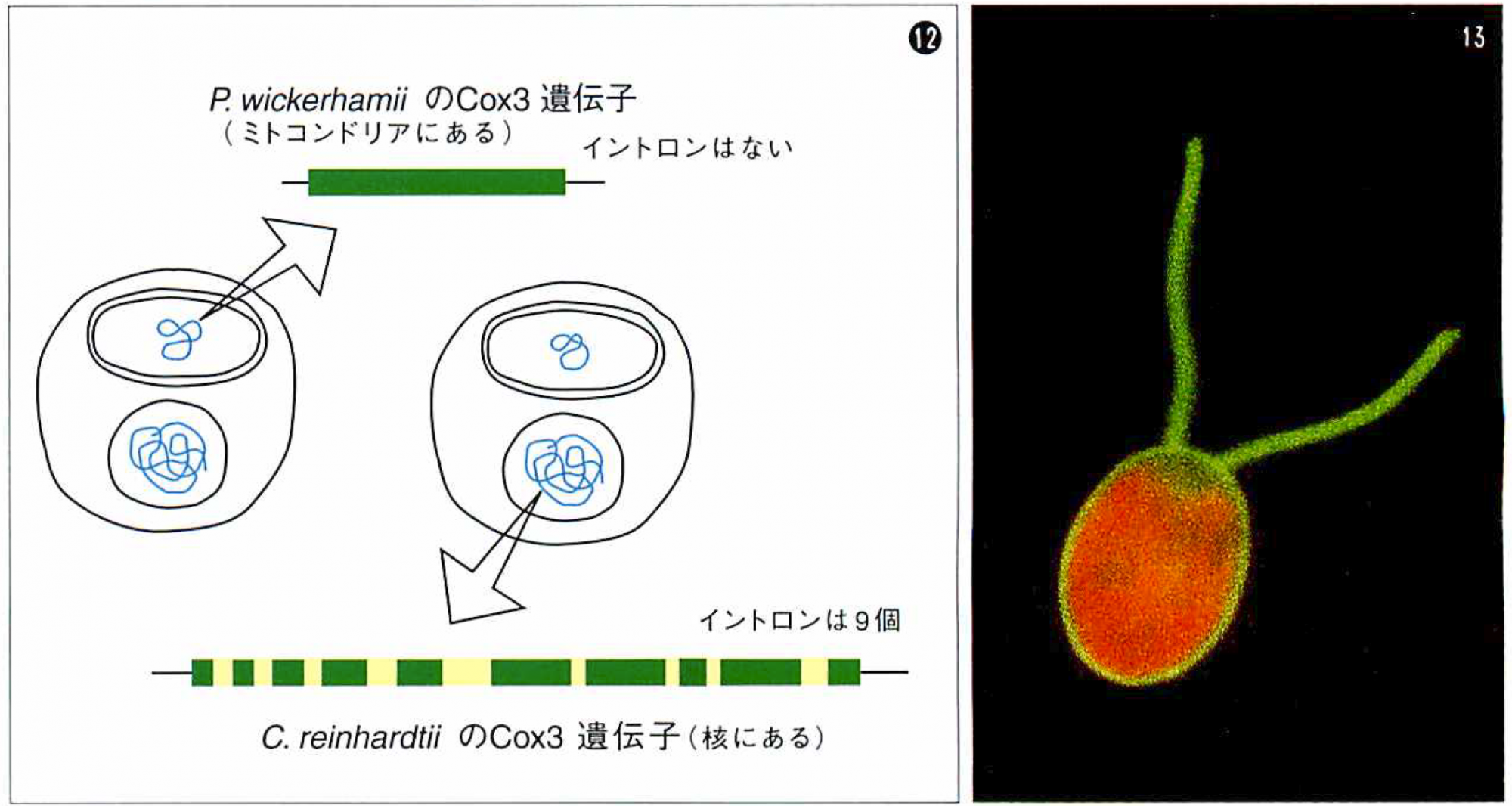
核型イントロンがそれまでイントロンをもたなかった核の遺伝子に入り込んだ証拠
⑫クラミドモナスC.reinhardtiiのCox3遺伝子は,今から1~2億年ほどの期間にミトコンドリアから核に移動した。移動前はイントロンをもたなかったのが,移動してから現在までに9個ものイントロンが入り込んだと考えられる。1~2億年前にクラミドモナスと系統が分かれたP.wickerhamiiのCox3遺伝子(ミトコンドリアにある)にはイントロンがない。
⑬クラミドモナス。微小管という細胞骨格の一種を蛍光染色したもの。鞭毛がはっきり見える。(写真=ヴァージニア大学・R.A.Bloodgood)
核型イントロンは,この侵入という属性を今でももっていることを,大学院生の渡辺らが,単細胞緑藻のクラミドモナスの解析から示した。クラミドモナスでは,本来ミトコンドリアにあるべき電子伝達系の酵素(エネルギー生産に関わる)であるCox2とCox3が,1~2億年ほど前に核に転移したことがわかっている。この遺伝子に何と全部で15個にもおよぶ核型イントロンが挿入されていることがわかったのだ(図⑬)。つまり,1~2億年という短期間(進化では)に,これだけ多くのイントロンが侵入したということだ。程度の差はあるにしろ,おそらく他の生物種でもこれと同じことは起こっているだろう。
イントロンは今でも入ったり出たりしている
エキソン1Kbあたりのイントロンの頻度は,生物種によってずいぶん違う(図⑭)。どうやらこれは,種によって核型イントロンの侵入および増殖の活性が違うことの反映らしい。イントロンは特別な場合を除き,生物にとって厄介者である。たとえば,切り出しに失敗すれば命取りになりかねない(現に,スプライシング異常による遺伝病が知られている)。それでも,イントロンはゲノムから消えることはない。どうやら,核型イントロンの数は,新たな侵入と削除の平衡関係が決定しているらしい。

生物グループごろにイントロンをもつ頻度は異なる
⑭エキソン1Kb(1000塩基対)あたりにイントロンが何個入っているかを示した図。イントロンが多いか少ないかは生物の系統関係とは無関係だということがわかる。
(Current Opinion in Cenetics and Development, vol.8, p.638, 1998の図を改変)
ところで,イントロンの削除は,最初に紹介した成熟したmRNAがDNAへ変換(逆転写)され,ゲノムに組み込まれることで起こるらしい。そうであれば,多細胞生物の場合,生殖細胞で発現する,つまりそこでmRNAが作られる遺伝子しかイントロンはなくならないことになる。多くの遺伝子は,脳,肺,皮膚というように,生殖器官でない特定の器官でだけ発現するので,これら遺伝子にはイントロンが侵入することはあっても,イントロンがなくなることはないかもしれない。こうした視点から,多細胞生物の登場とゲノムの進化の関係を考えると興味深い。
生物のゲノムの中には,繰り返し配列(たとえばCGAという並びが繰り返し,その数によってハンチントン病になるなどの例がある)や,トランスポゾン(動く遺伝子で薬剤耐性を運んだりする例がある)のような多くの利己的なヌクレオチド配列が含まれている。こうした配列は増えることだけに意味があるとしか思えないのだが,核型イントロンもそうした利己的な因子の一種であり,真核生物誕生のための負の遺産と見るのが,その本質を捕らえた,生命誌の視点からの理解に思える。イントロンが一部機能を担うようになったのは,二次的現象である。ゲノムにあるものすべてが,役をなすとは限らない。生物の隣には,必ず生物がいる。人間の歴史と同様に他を侵略し懐柔しながら生物は生き長らえている。ゲノムも,一つの個体,一つの種に注目するだけでなく,生物の相互干渉に注目すべき時が来ていると思う。
[あとがき]
大濱がJT生命誌研究館を仕事場とした,1994年から2000年のうちの後半は,藻類のミトコンドリアに見られるイントロンの種間転移の解析が主なテーマでした。江原恵・渡辺一生(かずお)の両博士(現日本学術振興会特別研究員)には,大阪大学理学研究科の博士課程院生として,このテーマで実験をしてもらいました。“どうして,イントロンはなくならないのかしら?”と大切な問題を再認識させてくれた中村桂子副館長,直接,間接的に研究をサポートしてくれた研究仲間や事務,研究補助員の方々に改めてお礼申し上げます。また,プロトシフォンでgroupIイントロンを見つけ,この研究のきっかけをつくってくれた石丸八寿子さん(現未来開拓特別研究員),ポスドクとして研究に参加してくれた平岩呂子さん(現株式会社アピ研究員)に感謝します。
(おおはま・たけし/JT生命誌研究館主任研究員・現高知工科大学物質環境システム工学科教授)
※所属などはすべて季刊「生命誌」掲載当時の情報です。













