最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。
バックナンバー
【収斂進化2:真獣類の大陸内適応放散と収斂進化】
2005年2月15日
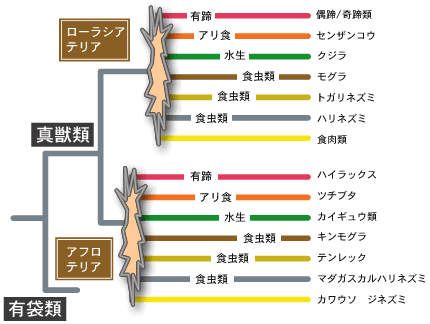
食虫類は非常に古い形態を示す系統を含んでいることが昔から知られていて、一つのグループにまとまらず、いくつかの独立なグループ(亜目)に分類されていた。ほ乳類の目に対応するグループは短期間に爆発的に多様化したため、厳密な分子系統樹の構築は非常に難しい問題である。スプリンガーらは、3つのミトコンドリア遺伝子と2つの核にコードされている遺伝子を使って真獣類の系統樹を推定した。その結果は予想通り食虫類は一つのグループにまとまらなかった。興味あることに、アフリカの食虫類の一つキンモグラはモグラやハリネズミよりも、むしろイワダヌキ、マナティー、ゾウ、ハネジネズミ、ツチブタといったアフリカ起源のグループに近縁であるということが示された。このことから彼らは、アフリカにおいて適応放散によってアフリカ固有の真獣類のグループ(アフロテリアという)が生じたと考えた。スプリンガーらとオブライエンらは解析をさらに進め、アフロテリア以外に、ローラシア大陸(北米大陸とユーラシア大陸から成る古生代末期の超大陸)固有のグループ(ローラシアテリア;クジラ、ウマ、ネコ、コウモリ、モグラ、などを含む)の存在を見つけた。それ以外にも南米の固有のグループ(ゼナースラ;ナマケモノ、アリクイ、アルマジェロを含む)、及びユーアーコンタ&グリレス(カピバラ、モルモット、マウス、シマリス、ナキウサギ、等を含む)の存在を見つけ、真獣類は大きく4つのグループに分かれて、それぞれが大放散したことを示した。日本では長谷川政美が早くからこの問題に取り組んでいて、幾つかの成果を上げている。 この結果は現時点ではまだ仮説に域を出ていない。今後多くの遺伝子とより厳密な解析によって確認されねばならないが、作業仮説としてきわめて興味深い重要な問題を含んでいる。スプリンガーらも述べているが、もしこの結果が正しいなら、大陸ごとに真獣類の適応放散が起こり、大陸間で多くの収斂進化が起きたことになる(図1)。有袋類と真獣類の間では多くの収斂進化が認められるが、真獣類の内部でも大陸間で頻繁に収斂進化が起きたことになる。なぜ、それほど多くの収斂進化が起こりうるのか?収斂進化の分子機構はなにか?今後の分子進化学に残された大きな問題の一つであろう。 | |||||||
|
|||||||
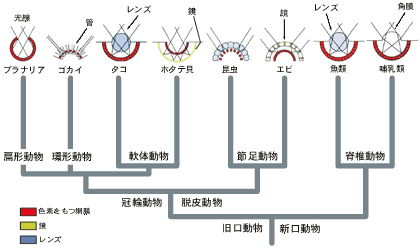
| 最後に最近の分子生物学からのデータをもとに眼の収斂進化を分子の視点で考察してみよう。眼は動物の行動にとって非常に重要な器官である。そのため、さまざまな動物の系統で多様な眼が進化した(図2)。われわれヒトを含めた脊椎動物の眼はカメラに似た構造を持っていることからカメラ眼と呼ばれている。カメラ眼は脊椎動物に限らず、軟体動物のイカなどにも見られる。但し、両者の間では網膜と視神経の位置関係が逆転していて、収斂進化の典型的な例としてしばしば教科書に登場する。 | |||||||
|
|||||||
|
最近、眼の形態形成遺伝子であるPax6がショウジョウバエとマウスから単離され、その後いくつかの動物から相同な遺伝子が相次いで見つかっている。さらにショウジョウバエの実験から、この遺伝子を異所的に発現させると、本来あるべき位置とは異なった場所に眼ができることが明らかにされている。従ってPax6は発生の過程で将来眼になる位置を決定する役割を持つと考えられる。また、軟体動物と節足動物のPax6を入れ替えても眼が形成されることも分かっている。こうしたことからPax6は眼の形成に関するマスター制御遺伝子であると考えられている。 さてPax6はマウスのカメラ眼やショウジョウバエの複眼のみならず、最も原始的な眼とされるプラナリアの眼にも相同な遺伝子が存在していることが明らかになってきた。Pax6遺伝子が眼を持つ動物に共通に存在することから、ウォルター・ゲーリングは脊椎動物と軟体動物のカメラ眼のみならず、昆虫の複眼も含めて、あらゆる動物の眼は収斂進化ではなく、共通の祖先から分岐進化(Divergence)によって多様化したと考えている。 なるほどPax6だけを見ていると眼は分岐進化に見える。しかし、今問題にしているのは、脊椎動物のカメラ眼と軟体動物のカメラ眼は機能・構造的に似ており、こうした眼の部分的類似性が収斂的に進化したのかどうかという点であって、眼という器官全体の進化パターンを問題にしているのではない。カメラ眼を持つ軟体動物のPax6を複眼を持つ節足動物のPax6と取り替えても眼が生じるが、いぜん複眼ができるのであって、カメラ眼が生じるわけではない。このことはPax6によって将来眼になる位置が決定されるが、複眼やカメラ眼といった眼の構造を決定するのは他の遺伝子(群)ということになる。 たしかに眼の形成に関与する制御遺伝子はPax6だけではない。Pax6の後にカスケード的に働く幾つもの制御遺伝子のセットが存在することが知られている。従って問題は、カメラ眼の形成に直接関与する遺伝子群がカメラ眼を持つ脊椎動物と軟体動物の間で相同なのか、あるいは全く由来の違う遺伝子セットなのかということになる。前者であれば分岐進化の可能性が大きいであろうし、後者なら収斂進化ということになろう。もちろん前者であっても遺伝子のセットが完全に相同というわけではなく、一方の系統では部分的にある遺伝子が欠失したり、別の遺伝子が付け加わったりすることもあろう。現在、ショウジョウバエやマウスでは眼の形成に関与するPax6以外の遺伝子が少しずつ明らかになってきているが、両者でそれらが互いに相同であるかどうか必ずしも明らかでない。こうした視点から近い将来形態レベルでの収斂進化を遺伝子レベルから捉え直すことができるようになると期待している。 眼は動物の行動を支配する重要な器官であり、そのため動物進化の過程で自然選択が強く働いたと思われる。まだ想像の域を出ないが、新しい環境に遭遇した動物たちは、速やかに与えられた問題を解決するために手近な「遺伝子」を利用してさまざまな眼を進化させていったのではないだろうか。こうした便宜主義が遺伝子レベルでも働いているなら、眼の遺伝子セットは動物の眼ごとにずいぶん違っているのではないだろうか。使っている遺伝子セットがかなり違っていても、表現型では一部を除けば非常によく似た構造(あるいは機能)を持った眼(例えば脊椎動物と軟体動物のカメラ眼)が進化することもあるかも知れない。 カンブリア紀と先カンブリア時代の境に起きた三胚葉動物の未曾有の多様化は、新たに新規遺伝子を作ることなく、多細胞動物の進化のごく初期に既に存在していた遺伝子を利用して達成したことが明らかになっている(本シリーズ第一回参照)。すなわち、この大進化においてさえも「便宜主義」なのである。生物は、多少出来上がりが不完全なものであっても、手近にある既存の遺伝子を利用して速やかに与えられた問題の解決を図ってきたのであろうか。かつてわが国が生んだ偉大な分子進化学者、木村資生は「表現型レベルの進化は便宜主義、分子レベルの進化は保守主義」と言っていた。これはあくまでも筆者の私見だが、これまでのわれわれの多様性研究の結果からすると、いずれのレベルでも便宜主義が横行しているようだ。 | |||||||
|
[宮田 隆] |