顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
デニソーバ人からの贈り物
2014年9月16日
14話のチベット族が高地順応して来た歴史を再度まとめておく。
「少なくとも1万年以上前にチベット族の先祖が高地へ移り始める。最初は私たちが高地に順応するのと同じような低酸素反応で高地に対応していたはずだ。しかし徐々に高地生活に有利な遺伝的変化が選択され、心肺機能、代謝機能等々を高地向きに変化させて行く。そして8000年ほど前に、低酸素反応の閾値を下げるEGLN1突然変異が起こり、高地でも赤血球やヘモグロビンが低いまま維持されるチベット族の性質が完成する。この過程でチベット族の先祖と同じように高地に住むシェルパの先祖の交雑が起こっている。おそらく、交雑は有利な遺伝子を一度に受け継げる大きなチャンスとなったと思われる。」
これが今想像されているシナリオだが、最初にチベット族のゲノムについて報告した2論文のうちの一編では高地順応との相関性が最も高い多型としてEPAS1遺伝子領域の多型について指摘していた。このグループはその後もこの遺伝子領域を詳しく検討し続けた。EPAS1はHIF2αとも呼ばれ、HIF1のように低酸素に反応して血管や心臓の働きを高め、赤血球を増やす働きがある事がわかっている。EPAS1遺伝子領域を含む32000塩基対の長さを持つ領域の中に、明らかに高地順応に関わると思われる多型(チベット族だけに見られる印)がまとまって存在する。祖先が同じ民族同士でも時間がたてば当然多型が生まれるのだが、これほど小さな領域にまとまって多型が片方の民族にだけ存在する事はまずない。高地順応過程で独立に生じた多型が積み重なったとも考えられるが、様々な計算を行うと、これだけの数の違いが別々に選択される事は考えにくい事がわかった。従って、先祖が他の民族との交雑によってこの領域がそっくりチベット族に移って来た後、この領域を持つ人達が選択されたと考える可能性が高いと考えられた。これが正しいとすると、世界中を探せばこの領域がチベット族と同じタイプの多型を示す民族がいるはずだ。この仮説に基づいて、このグループはこれまでこの部位の塩基配列がわかっている民族をしらみつぶしに探した。勿論一つ、二つの多型がチベット族と共通の民族はいるにはいたが、これだけまとまって多型がかたまっている民族を見つける事は出来なかった。ところが探索を既に滅びた人類、ネアンデルタール人やデニソーバ人に拡げた所、驚くなかれ、デニソーバ人のこの領域が、チベット族とほぼ一致する事がわかった。この32000塩基対の長さの領域だけでチベット人と、デニソーバ人を含む様々な人種を比較すると、やはりデニソーバ人が最も遺伝的に近いと言う結果が出る。14話で紹介した様に、シェルパとチベット族との間で交雑があった事から考えると、チベット族かあるいはシェルパ族の先祖が何万年も前にデニソーバ人と交雑することでこの遺伝子領域がチベット族の先祖に流入したと考えるのが最も可能性が高い。面白い事に、やはりデニソーバ人から流入して来た遺伝子を持つ事がわかっている、メラネシア人ではこの領域は現代人型だ。従って、チベット族の先祖が高地に移り住む事でこの領域が有利に働き、チベット族全体に拡がったが、平地に住む集団からは消えて行ったと考えられる(図1)。(Nature, 434,288-289, 2014)
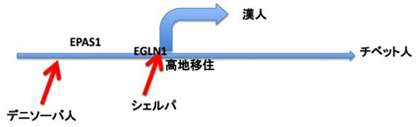
図1:チベット族の歴史:チベット族が高地順応する過程でEPAS1,EGLN1遺伝子多型を獲得するが、EPAS1領域の多型はデニソーバ人由来である事がわかった。(Nature, 434,288-289, 2014)他にもシェルパ族との交雑も確認されており(Nature communications,5, article No. 3281, 2014)、集団内の変異だけでなく、交雑を通した遺伝子流入で高地順応した事がわかる。
しかし、デニソーバ人とは何者か?なぜ絶滅した原人のゲノムが解読できているのか?この問題に私たちが答える事が出来るのは、ドイツライプチッヒにあるマックス・プランク人類進化学研究所のスバンテ・ペーボ(Svante Pääbo)さんたちの研究による所が大きい。彼を一躍有名にしたネアンデルタール人についての詳しい紹介は次回に行うので、今回はデニソーバ人とは誰かについてのみ紹介する。ペーボさんたちによって、化石から遺伝子が回収できる事がわかってから、保存のいい骨の化石の持つ価値が認識されるようになり、よく保存された骨が欧州各地で探索されるようになった。ただ、直立原人(エレクトゥス)以前の化石はどうしても温度の高い領域にあるため保存状態が悪い。この問題を解決できる格好の発掘場所として注目されて来たのがアルタイ山脈だ(図2)。

図2 アルタイ山脈とデニソーバ洞窟。既に観光客が訪れる名所になっている。(Wikimedia Commonsより)
ここは緯度が高く、高地にあるため化石の保存状態がいい。残念ながらここから出土する骨は何故か完全な骨格が失われており、歯や骨の断片だけが出土していた。しかし期待通りDNAの保存状態がいい事がわかり、2008年にこの地域にあるデニソーバと呼ばれる洞窟の3−4.8万年前の地層から出土した指の骨からDNAが抽出され、ゲノム解析が始まった。まずこのゲノムに含まれるミトコンドリアDNAの全配列が決められた。(Nature, 464,894, 2014)。初めはネアンデルタール人の骨かと考えられていたが、なんとネアンデルタールでもない、ホモサピエンスでもない新しい類人を代表するゲノムである事が明らかになった(図3)
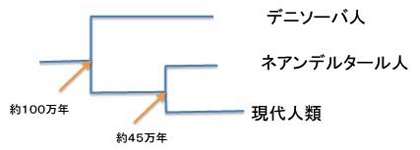
図3:ミトコンドリアゲノムから推定される3類人の関係。(Nature, 464,894, 2014)
その後このDNAを用いてデニソーバ人の全ゲノム配列が決定された。この配列データを比べる事で、デニソーバ、ネアンデルタール、現代人がどこかで出会い、交雑したかどうかを推察する事が出来る。先ず、デニソーバ人の遺伝子配列には0.5%程度のネアンデルタール遺伝子の流入が認められる。従って、両方はどこか出会い、交雑が行われた事は確かだ。しかしデニソーバ人とヨーロッパ、アフリカ、アメリカに現在住んでいる現代人類とはほとんど遺伝子の流入がないと言っていい。ようやく、アジア人のゲノムには0.2%程度のデニソーバ人遺伝子の流入痕跡が見つかる。驚く事にポリネシアやオーストラリアの原住民の遺伝子の3−6%もの遺伝子がデニソーバ人由来である事がわかった。この事から、デニソーバ人と現代人の一部が出会って交雑を行っていることはたしかだ。デニソーバ人の遺伝子がポリネシアやオーストラリアの原住民にしか見られない事から、交雑はかなり限られた集団とだけ行われ、最終的にその集団はポリネシアへ移住し、他の現代人の集団から隔離されていたおかげでデニソーバ遺伝子を多く残したまま現在に至っていると考えられていた。そこに今回、チベット人のEPAS1遺伝子領域が、デニソーバ人とほとんどそっくり同じだとするこの論文が現れたから大騒ぎだ。残念ながらデニソーバ人が高地順応していたのか、どの程度の期間現代人と同じ領域に共存していたのかなどまだまだ謎は多い。もっと多くのデニソーバ人のゲノムが解析されないと、実際にどんな事が起こっていたのか推論する事はまだまだ難しい。しかしこれも時間の問題と確信できる。この確信の背景には最近急速に進んだネアンデルタール人のゲノム研究の成果がある。次回はこれについて紹介する。