顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
III 新しい情報社会
2019年3月1日

これまで見て来たように、文字は出版と一体化することでそれまで一部のエリートに独占されていた文化を大衆化する原動力として働いてきた。文字自体はアルファベットが発明され、様々な言語で使いやすく改良されて以来、特に大きな革新はなくなったが、様々な出版の技術革新が起こるたびに大衆化は促進する。中でももっとも大きな転換点になったのがグーテンベルグの印刷術の発明だった。言い換えると出版が機械化され、機械の効率を上げれば出版の効率を高められるというサイクルが完成した。この機械化は出版の中央集権的組織化のを促し、その後出版はビジネスとして利用者を拡大し、そのまま20世紀に至っている。特に20世紀に入って。出版も写真などの新しい情報メディアが取り込まれることで、出版は完全に大衆文化の中心に躍り出る。
しかし、20世紀が終わり21世紀に入るとすこし様相が変わってくる。出版協会が発表している日本の出版販売額の推移を見ると(図1)、21世紀に入る頃からの販売額は書籍も、雑誌も右肩下がりになっている。
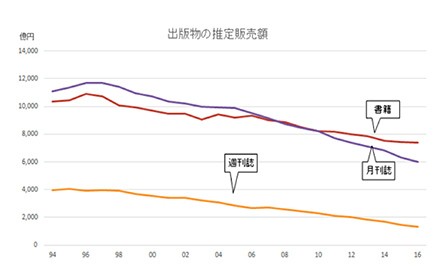
新しい本の出版数はそれほど落ちていないようなので、この統計から本を買って読む人の数が減っていることがわかる。この最大の原因は、個人と文化の接点としての書籍の位置が低下してきたことによるだろう。
勿論この変化の背景の1つは、文字に変わる様々なバーチャルメディアの発展がある。文字にせよ、絵にせよ、人間が主観的に書き残したものの最大の問題は信頼性の問題だ。例えば、公的な歴史は捏造されていることが多い。支配者が少しでも格好良く見せたいと思うのは当然のことで、政府プロパガンダや反政府プロパガンダに凝り固まった歴史家は別として、普通の歴史家は起こった歴史的事件の内容を読み解くため、1つの記録をそのまま信じることはない。政府による捏造は専制国家だけの話かと思っていたら、昨年我が国で政府の見解に合わせて多くの公的文書が改ざんされたことが明らかになった。恥ずべき話だが、我が国のような民主国家でさえ、書くという主観的行為に内在する難しい問題によって蝕まれていることを思い知らせてくれた。一般的に、その事件の当事者の記録は誇張や捏造が多く、利害のない観察者の記録は比較的信頼性がある。これを読まれている皆さんも、自分のことを書くとき、捏造しないまでも良い格好をしたり、誇張したりしていることに思い至らないだろうか。
この問題を解決するには、経験者が主観的な言葉や絵として書き残すのではなく、出来るだけ客観的なメディアを利用することが重要になる。この真実を記録する鏡を可能にした最初の技術的成果が写真ではないだろうか。カメラが映し出す景色は捏造や嘘のない世界が切り取られていると多くの人が期待を持った。この期待を受けて、写真は書かれたことの信頼性を高める役割を果たし、出版による文化の大衆化をさらに促進し、1920年以降、報道写真という一分野が確立する。
しかし、もちろん写真も捏造や脚色から解放されているわけではない。実際、報道写真や映画を個人的考えに沿った形で編集できることは明らかで、現在でもこういった捏造を報道から根絶するためには、複数の記者が同じ場面を記録する以外に方法がなく、基本的には報道する側の倫理観を信頼する他ない。
逆に写真を使って主観的な世界を表現した虚像を、実像と錯覚させられる可能性は、多くの芸術家の想像力を刺激し、虚構の世界を表現する芸術の世界で写真技術が用いられるようになる。この先駆けになったのが、シュールレアリズムの表現手段として写真を利用したマン・レイだろう。この虚構と現実の緊張感をさらに発展したのが今私達が映画やテレビで見ている、多くの映像娯楽作品と言える。

もちろん虚構の世界を表現するだけでなく、写真、録音、映画、ビデオなど新しいメディアの全ては、実像を切り取りコピーするという本来の能力を急速に高めてきた。客観的な事実の記録が要求される科学研究が、今やこれらの記録技術なしに存在し得ないことは、この進展を示す良い例だ。同じように、事実を報道するという目的についても、報道側の倫理観が重要であるという前提はあるものの、Writingに頼らない報道のための新しいメディア技術の発展は、20世紀文明を特徴づける進歩の一つだろう。
大事な事は、この新しいメディア技術の普及が2段階で進んだ点だ。まず放送局という新しい中央集権的マスメディアが誕生し、情報を効率よく組織化して伝える技術を発展させる。これが、おそらく世界中の人が一日のかなりの時間見たり聞いたりして過ごすマスメディアだ。これまで、この放送局を中心とするメディア、特にテレビの発展は、ともするとWritingを接点とする大衆文化を破壊すると考えられて来た。グーテンベルク以来発展を続けてきたWritingをベースにした大衆文化をテレビが破壊することを懸念した言葉が「一億総白痴化」だ。しかし我が国で言えば1953年の公共テレビ放送開始以来、出版点数は右肩上がりで2000年まで増加の一途をたどっている。当時1−2万点数の間だった出版点数は、2000年には70000点近くと、3倍以上に増えている。技術革新で本を作るのが簡単になったこともあると思うが、これらの統計を見ると、テレビと活字媒体はうまく領域を分け合って、共に繁栄して来たと言えるように思う。
テレビやラジオではあまり努力することなく、自然に情報がインプットされるが、逆に情報を知るためには一定時間メディアに向き合い続ける必要がある。一方活字媒体では、主体的に情報を選ぶことが出来るし、瞬時に内容を理解できる短いサマリーで表現することが出来る。このように、Writingにはテレビやラジオにはない素晴らしい特徴が多く存在している。このためどんなに大衆と知的エリートの間のギャップが拡大した社会でも、文字を媒体にした情報メディアが廃れたわけではない。このことは、タブレット版の大衆紙が発行部数は落ちたとはいえ、世界中で今も健在であることからよくわかる。
しかし21世紀に入ると、出版をベースにした大衆文化は大きな変革の波にさらされ始める。この大きな要因が、メディア技術革新の第二段階、すなわち見る、聞く、読むに加えてコミュニケーションの全ての手段が一体化した新しい個人ツールが開発されたことだ。スマートフォンを所有している人は、使う、使わないに関わらず、Writing、写真、録音、ビデオを所有していることになる。面白いのは中央集権的放送の第一段階から個人が中心の第二段階へと進むこの順序が、出版が中央集権的に組織化され、大衆化した過程の逆をいっている点だ。すなわち出版は、個人のWritingを拾い集め、コピーして大衆に流通させる中央集権的システムといえるが、これが実現するには印刷の機械化が必要で、これは出版の歴史でかなり後に実現した。この機械化により出版はより中央集権化し、これによる効率の上昇に合わせて、大衆化した。
一方、写真やビデオといった新しいメディアについても根本の構造は同じなのだが、これらのツールが発明された当初は、機器の価格が高額で個人が普通に手を出せるものではなかった。このため、情報の収集と提供は最初から放送という中央集権的システムから始まらざるを得ない。個人は、レシーバーだけを持ち中央からの情報提供を待つ。商業放送では、今もこの形式は変わっていない。いずれにせよ、情報の流れから見ると、現在の出版も放送も同じような中央集権化した階層性でできている(図3)。
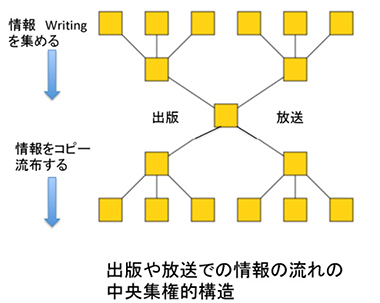
図3 放送、出版での情報の流れ。
手紙のように個人のwritingは中央集権的ではないが、出版というプロセスはコピーしそれを大勢に届ける必要性に応えるため、必然的に中央集権的になる。歴史的にはこの中央集権的構造はグーテンベルグの印刷術の発明で確立する。一方、放送に関しては、最初から中央集権的にならざるを得なかった。
ところが21世紀、放送や出版に必要なこの構造が大きく変わり始めた。情報を記録するツールがほぼ全ての個人に行き渡り、それを多くの人に提供できるITネットワークを誰もが利用するようになった。スマートフォンにWriting、写真、録音、ビデオと放送局顔負けのツールが備わっていることも重要だが、それ以上に重要なのはそれぞれの端末とそれを持つ個人がネットで繋がっているという事実だ。これをPeer to Peer(P2P)ネットワークと呼んでいるが(図4)、このおかげで一人の個人的体験を瞬時に端末を持つ全ての人に伝えられる構造が完成した。そしてこの誰もが瞬時に結ばれるP2P構造が既存の中央集権的体制を蝕み始めている。
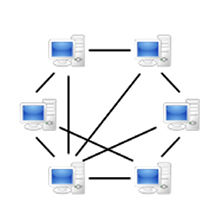
図4 Peer to Peerネットワーク構造。発信と受信が可能な双方向的端末が、多くの端末と直接、間接的に連結している。
残念ながらこの構造は決して自発的にボトムから出来上がったわけではなく、個人がつながる為に特定のアプリが必要になる。例えばFacebook, Twitterに始まり、YouTube、我が国ではニコ動、LINEなどにより提供されるプラットフォームが必要だ。例えばFacebookは決してLINEと相互乗り入れできない。ビジネス的に言うと、コンテンツを提供せずに、情報がやりとりできるプラットフォームを提供するだけのビジネスが誕生した。このようなネットワークを整備すれば、コンテンツはそこに自発的に集まり流通できる。これらの企業が21世紀を代表する大企業に育ったということは、P2P構造が社会構造を変えつつあることを示している。しかし個人の立場から考えると、プラットフォームを一定の企業が独占するということは様々な問題を伴う。下手をすると、ジョージ・オーウェルの小説『1984年』に出てくるビッグブラザーを誕生させる危険すらある。従って、将来はこのP2Pを支えるプラットフォーム自体のあり方も変わるような気がする。いや、変えるように努力すべきだろう。
とはいえ、P2Pネットワークの場合、複数の人が目撃し記録をとっておれば、個人的な捏造の動機を完全に排除できる。例えば、複数の人が同じ画像をスマートフォンで写して拡散すれば、皆で示し合わさない限り捏造は排除できる。これにより中央集権的な情報の流れが完全に壊れたことを意味している。
面白いことに、この全く新しい可能性は、既存のメディアを否応なくSNSの積極的ユーザーへと駆り立てた。スマートフォンによる個人の情報蒐集網は世界の片隅張り巡らされており、既存のメディアもこの魅力に屈するのは当然だ。この結果、既存のメディアがSNSと競争して外部から侵食されるだけで無く、麻薬のように内部に浸透したSNSにより侵食されるようになった。
これは報道だけでない。様々なメディアを用いたフィクションも、同じネットを用いて流されるようになる。映画、CD、ビデオの代わりに、YouTubeなどの映像配信プラットフォームの躍進は著しい。文学についてはまだまだ出版の役割は大きいが、それでもウェブで自由に小説を発表できることは間違いない。スマートフォンとの相性のいい漫画では、すでに漫画作品の投稿サイトがLINEなどにより提供されている。
こうしてみてくると、言語コミュニケーションが数万年前に始まって以来、できるだけ多くの人とのコミュニケーションを求めて発展を続けた結果、個人が瞬時に世界中の人とコミュニケーションできる現実が21世紀に完成しようとしているのがわかる。この先にどんな未来があるのか?
これまで言語の誕生、Writingの歴史を概観してきたが、ここで得た知識をもとに考えてみると、おそらくアルファベットの発明とともにギリシャで起こった、新しい大衆文化の発展に匹敵する変革が21世紀に起こる予感がする。現状だけを見ると新しい可能性に様々な問題は存在するが、これらは全て過渡期の現象だと私は思っている。そして21世紀中には、中央集権的な放送や出版はなくなるとすら思っているが、それに代わって何が残るのかは想像がつかない。ギリシャはこの時民主主義を発展させた。まさに同じ直接民主主義を可能にするプラットフォームは完成しつつある。しかし最初の民主主義を経験したプラトンが著作『共和国』で述べているように、自由を善として発展した直接民主主義は自ら滅び、優れた指導者による僭主政治を待望する時代がまたやってくるのかもしれない。
予告
2019年3月末で私は生命誌研究館を去ることになり、「進化研究を覗く」もちょうどキリがいいので、これで終わりにする。しかし研究館は辞めても、私自身は別の場所で様々な活動を続けていく予定で、このコラムはそのまま私のNPOのホームページ(図1)へ引っ越すことにし、準備を進めている。
図1 AASJホームページ。このコラムの続きも含め、様々な企画が新しく始まるので、ページの構成は変わると思うが、www.aasj.jpのアドレスで全てのコンテンツを自由に閲覧することができる。4月1日には以下のコンテンツもスタートできるよう努力している。
新しく企画しているのは何度も予告したように「生命科学の眼で読む哲学書」というタイトルで、ギリシャから現代哲学までを、生命科学者の独断と偏見で紹介してみたいと思っている(図2)。私にとってこの作業は、生命科学がどう成立してきたのかを自分なりにまとめるための大事な作業だが、これまで生命科学について書かれているサイトとして読んでいただいた読者にとっては、あまり馴染みのない哲学者の話になってしまうと思う。

図2 現在計画中のコラムでAASJホームページから自由に閲覧できるようにする。
ただ「生命科学の眼で読む」ので、普通の哲学書紹介とは全く違った観点から紹介する。ぜひこの機会に、過去の哲学と現代の生命科学との関係について、一緒に考えて欲しいと思っている。
また、これまで研究館のHPに掲載してきた記事は、「生命と情報の誕生」、「脳活動から言語へ」、そして「Writingとマスコミュニケーション」としてまとめ直して、AASJで公開し直す予定なので、今後も記録として自由に閲覧、利用していただければ幸いだ。
最後になるが、2014年からほぼ5年にわたって、生命科学の過去、現在、未来をまとめるためのノートとしてこのコラムを書いてきた。これを支えていただいた、研究館のスタッフ、特に現在熊本にいる有馬さん、そして表現セクターの川名さんに謝意を表したい。
終わり。





