植物の上陸に適応した受精のしくみ
被子植物の繁栄を支える
重複受精の瞬間を見る
海で生まれた植物が上陸し、乾燥した陸地でも繁殖できるようになる過程で大きく変化したものの一つに受精のしくみがあります。コケやシダ、裸子植物までは精子は水中を遊泳し、鞭毛の遺伝子が存在しますが、被子植物ではそれがなくなります。花粉管が素早く伸び、精細胞を卵細胞に送るという陸上に適応した受精に変わるのです。このとき卵細胞の隣にある助細胞が誘引物質を出します。花粉管が取り込んだ2個の精細胞のうち、1つが卵細胞、もう1つは中央細胞と融合して、前者は胚、後者は胚乳になります(重複受精)。胚が常に胚乳という栄養分と共にあり、荒れ地でも発芽できることが被子植物の繁栄を支えました。名古屋大学の東山哲也さんは、生きたまま生殖細胞を観察する方法を工夫し重複受精のしくみを解明しています。
1.被子植物の誕生と繁栄
藻類として海に現れた植物は、今から約5億年前陸上に進出し、裸子植物から被子植物へと進化し、徐々に乾燥した土地へも広がっていった。現在最も繁栄している植物は、花を咲かせ実を結ぶ被子植物である。(図1)
藻類から裸子植物までの受精は、鞭毛や繊毛を備えた雄の配偶子が水の中を泳いで雌の配偶子に出会うというしくみがほとんどである。上陸後の植物にとっては、限られた水をいかに効率よく利用して受精するかは、種の存続をかけた最重要課題の一つだったに違いない。コケやシダの精子は、雨水など周りの環境から得られる水を頼りに卵細胞までたどり着く。イチョウでは、風で飛んできた花粉が、むき出しの胚珠(註1)の先端から出入りする液滴につかまって胚珠の中に引き込まれ、その後数ヶ月かけて胚珠内をゆっくり伸長する花粉管の中で2つの精子が形成される。花粉管から泳ぎ出た精子は、胚珠を満たす水の中を泳ぎ、卵細胞に到達して受精する。

(図1) 植物界の大まかな種数
被子植物は桁違いに種類が多い。(値は『数値でみる生物学』より引用)
しかし、被子植物になると精細胞の運動能力が失われ、新しい受精のしくみを獲得する。それは重複受精と呼ばれ、20万種を超える被子植物に共通のしくみである。重複受精は、水の少ない陸上環境に適応し、素早く種子形成するという特徴を持つ。砂漠に降った雨で種子がいっせいに芽吹き、花をつけ、水が干上がる前に次世代の種子をつくる様子は、まさに被子植物の適応能力の高さを示し、これが繁栄をもたらすと考えてよかろう。
2000年にシロイヌナズナのゲノムを解読したところ、鞭毛ダイニンの遺伝子は見いだせなかった。鞭毛を失った被子植物が新たに獲得した受精のしくみを見て行こう。
(註1) 胚珠
めしべの子房の中に作られ、後に種子となる器官。外側は厚い珠皮などの組織に覆われ、内部には卵細胞・中央細胞・助細胞などが納められている。ふつう胚珠には一カ所だけ孔(珠孔)が開いており、花粉管はそこを通って胚珠の中に進入する。トレニアでは卵細胞などが珠孔から外に突出している。
2.精細胞の運び屋:花粉管
被子植物の精細胞(註2)は、もはやイチョウの精子のように泳いで卵まで到達することはできない。そのかわり、時速数ミリメートルという植物としては非常な速さで花粉管を伸ばし、すばやく精細胞を胚珠に送り届ける。花粉管は、一個の花粉管細胞が細胞の一部を長く伸ばして作る構造である。花粉管細胞の中には、エンドサイトーシス(註3)によって生じた膜に包まれた二つの精細胞があり、花粉管が高速で伸びるのと一緒に先端へと運ばれていく。(図2)
このとき、雌しべの中で花粉管の伸びる方向を制御して、胚珠へと誘導するしくみを知ろうと古くからさまざまな誘引物質の探索が試みられてきたが、その実体は明らかにならなかった。
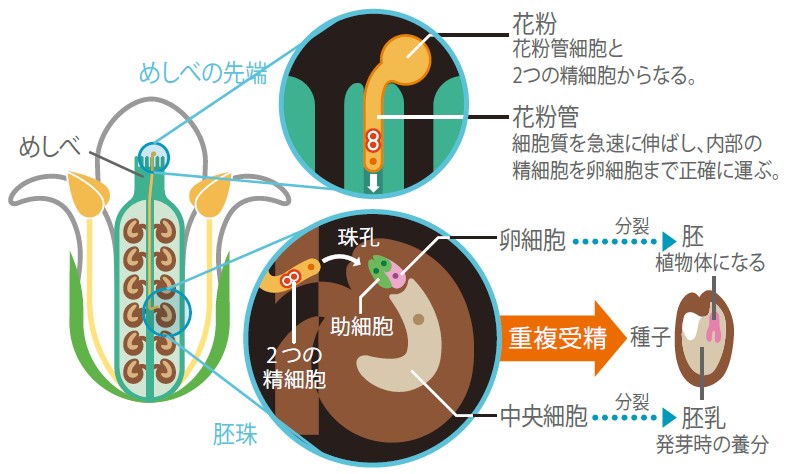
(図2) 雌花・胚珠の構造と、花粉管伸長の模式図
花粉管はめしべの中心を通り、胚珠まで精細胞を運ぶ。受精して発生が始まると、卵細胞は胚となり、中央細胞は胚乳となる。
(註2) 精細胞
配偶子の形態・機能に雌雄で著しい違いがあるとき、雄の配偶子のうち運動性を持つものを精子、持たないものを精細胞と呼ぶ。
(註3) エンドサイトーシス
細胞が外界の物質や細胞などを、細胞の中に取り込む現象。細胞膜の一部が取り込もうとするものを徐々に取り囲み、次にそれが陥入し、次第にくびれて細胞膜から切り離され、最終的には膜に包まれた構造として細胞内に取り込む。飲食作用。
3.花粉管ガイダンス分子
私たちは、トレニア(キク亜綱アゼトウガラシ科Torenia fournieri)という植物を用いてその解析を進めた(図3)。私が所属していた黒岩常祥先生の研究室では、興味をもった現象の研究に最適の材料を選ぶというポリシーがあった。私は受精研究に最適の材料を探し、トレニアに出会った。植物の受精に関わる卵細胞・二つの助細胞・中央細胞の合計四つの細胞は通常胚珠組織で覆われている。トレニアではこれらの細胞が胚珠組織の外に突き出ているので、生きたまま受精を観察し操作できると考えたのだ。(図4)

(図3) トレニア(Torenia fournieri)
園芸分野でも知られる身近な花。花粉管誘引ペプチドの発見に大きく貢献した。
大学院生時代にトレニアの体外受精系を立ち上げ、その後、高エネルギーの紫外線レーザーによって胚珠の細胞を一つ一つ壊し、花粉管を誘引している細胞を探った。その結果、卵細胞の隣にある二つの助細胞がその役割をしていることがわかった。
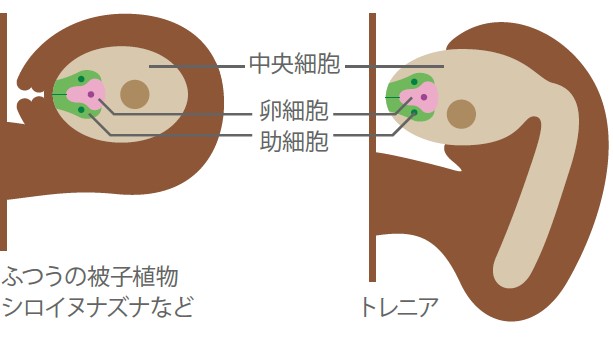
(図4) 胚珠の構造
トレニアでは生殖に大切な細胞がむき出しになっている。
そこで顕微鏡下で助細胞を取り出して遺伝子発現を調べると、驚いたことに多数の分泌性ペプチドが高レベルで発現していた。 その中に、LUREと名付けた花粉管誘引ペプチドを複数発見できた(図5、映像1)。
LUREペプチドは分子進化が速く、近縁種間でも配列が異なる。そこで同属別種のトレニアに対しては誘引が弱く、同科のアゼトウガラシではほとんど誘引が起きなかった。異種の花粉で受粉すると、近縁な植物の花粉管ほど卵細胞の近くまで進入する。LUREペプチドはこの最終段階ではたらき近縁な異種でも、交雑を起こさせないための重要な障壁としてはたらいていると考えられる。
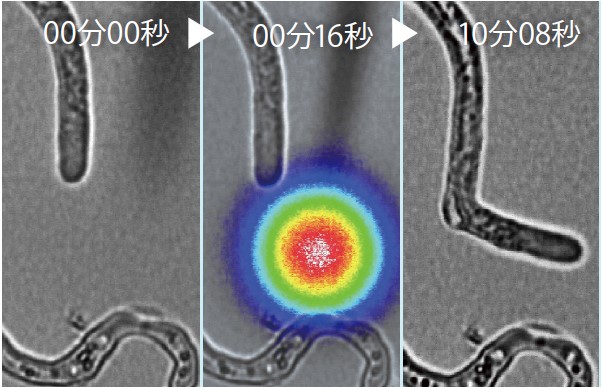
(図5) トレニアの花粉管がLUREペプチドに誘引される様子
虹色の同心円がLUREペプチド。色が赤いほど濃い。Okuda et al. (2009) Nature 458, 357-361 より転載。
(映像1) マイクロピペットで滴下したLUREペプチドへ向かって進む花粉管
Okuda et al. (2009) Nature 458, 357-361 より転載。
4.重複受精を捉える
胚珠で花粉管から放出された精細胞のうち、一方は卵細胞と、もう一方は中央細胞と受精する。受精すると、卵細胞は胚に、中央細胞は胚乳に成長する。しかし、めしべの奥深くで起こるこの現象を生きたまま観察することは難しい。
そこで、私たちはモデル植物シロイヌナズナを用いて遺伝子操作をし、生きたままでの重複受精の観察に挑戦した。このためには、①花粉管の中にある小さな精細胞など、受精に関わる雌雄の各細胞が光るシロイヌナズナづくり②高効率な体外受精法の確立③厚く覆われた胚珠内を撮影可能にするレーザー顕微鏡システムを整えた。
こうして、重複受精の瞬間を映像で捉えることに成功し、二つの精細胞がごく短時間で卵細胞と中央細胞のすき間に放出され、受精相手に到達する様子が観察できた。
二つの精細胞はおよそ7分ほどその場で留まった後、一方は卵細胞と、もう一方は中央細胞と受精する(映像2、図6)。受精の順番はきまっていないが、卵細胞と中央細胞の受精には約2分半の間隔がある。興味深いことに、卵細胞の受精に時間がかかったときには中央細胞の受精にも時間がかかるというように、二つの受精の時間には相関がある。二光子レーザー顕微鏡を用いて胚珠の奥深くをさらに詳しく観察すると、細胞のすき間に留まるあいだ、精細胞はただ細胞間隙に浮いているのではなく、卵細胞・中央細胞の両方と接した状態にあるように見えた。重複受精に欠かせない細胞間コミュニケーションがあるのだろう。
(映像2) 被子植物が受精する瞬間
二つの精細胞は花粉管から放出され、卵細胞、中央細胞と融合する。
撮影:浜村有希 博士(名古屋大学GCOEライブイメージングセンター・チーフコーディネーター)
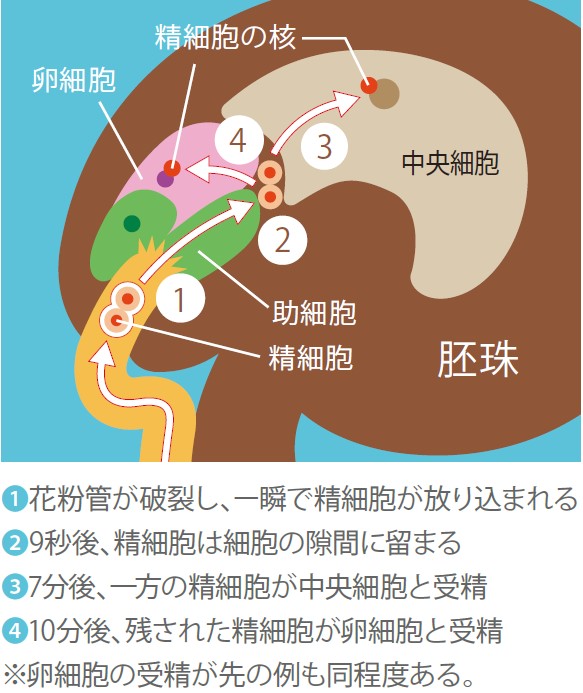
(図6) 受精の模式図
5.重複受精の謎
二つの精細胞が確実に異なる相手と受精できる理由については、これまで二つのモデルが唱えられてきた。一つは、精細胞の受精相手はあらかじめ決まってはいないが、多精拒否機構(註4)でもう一つの精細胞は未受精の細胞と受精するという考え、もう一つは、精細胞の受精相手はあらかじめ決まっているという考えである。
前者については、精細胞の数が増えるシロイヌナズナ変異体を用いると中央細胞だけで多精が起こるという報告があるので、卵細胞が先に受精し、残りの精細胞が中央細胞と受精する可能性が考えられた。ところが、両者の受精の間隔は予想よりはるかに短く、中央細胞が先に受精する場合もあった。おそらく卵細胞と中央細胞の両方で素早く多精のブロックがはたらき、しかもそれが局所的に起こるために精細胞の数が増えたときには巨大な中央細胞が多精しやすくなるのだろうと考えている。
後者のモデルについては、ナデシコ亜綱のルリマツリの仲間(Plumbago zeylanica)で有名な報告がある。この植物は、花粉管内で並ぶ二つの精細胞のうち後ろ側のものだけが色素体(註5)を持つので精細胞を区別できるのだが、後ろ側の精細胞の9割以上が卵細胞と受精したのである。
この植物のように二つの精細胞に明確な差が見られることは稀なのだが、花粉ができるときの二つの精細胞のでき方、並び方は被子植物に共通なので、一般に花粉管内の位置に応じて受精相手が決まっているとする説が長いあいだ支持されてきた。そこで私達は、紫外線を照射すると緑色から赤色に変わる蛍光タンパク質を用いて、シロイヌナズナで前後二つの精細胞を染め分けた(図7)。こうして、二つの精細胞の位置が花粉管内で入れかわることはないこと、前後どちらの精細胞も卵細胞とも中央細胞とも同程度に受精することがわかった。つまり、シロイヌナズナでは花粉管内での精細胞の位置と受精相手には関係がない。
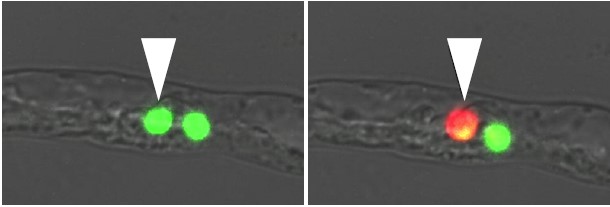
(図7) 染め分けた二つの精細胞
紫外線レーザーのスポット照射を行い、花粉管(右側が前)の中で、後方の精細胞だけを赤く染める。その受精先は精細胞の順序には関係なく、胚珠内での細胞間コミュニケーションで決まっている可能性が示された。
Hamamura et al. (2011) Curr Biol 21,497-502 より転載。
被子植物では、胚乳形成を促すよう雄型のインプリンティングを受けた精細胞のゲノムが中央細胞に持ち込まれることで胚乳形成が進む。二つの精細胞でインプリンティングの状態が異なるかどうかは不明である。しかしシロイヌナズナでは、両方の精細胞で胚発生にはたらく遺伝子が発現しており、片方だけで発現している遺伝子はこれまで見つかっていない。これらのことから私たちは二つの精細胞の機能は等しいのではないかと考えている。
実はイチョウの花粉にも精子は二つ入っているが、このうち受精に使われるのは一つだけで、イチョウの雌株は受精前から数ヶ月かけて、半数体の胚乳をゆっくりと成長させる。これに対して被子植物の重複受精では余った精細胞も利用して、効率良く胚乳形成を行えるようになった。私たちの実験はそんな可能性を示している。
一つ一つの細胞を顕微鏡下で自由自在に操り、オルガネラ、さらには分子に至るまでを解析したい。これが大学院生の頃からの夢であり、今、それが「ライブセル生物学」という分野として展開できるようになった。重複受精というテーマには、生きた細胞を操作して個々の細胞の役割を明らかにする手法を活かせる魅力がつまっている。夢を目指して研究を進めている。
(註4) 多精
一つの卵細胞に二つ以上の精細胞が受精すること。多くの生きものでは、一個目の精子が受精すると二個目以降の精子の受精を妨げるような反応が進むため、自然の状態では起こりにくい。
(註5)色素体
葉緑体、クロロフィルを欠いた白色体、デンプンを貯蔵するアミロプラストなど、植物細胞が持つ葉緑体の類縁オルガネラの総称。ミトコンドリアと同じく独自のDNAを持ち、その起源はシアノバクテリアの一種と考えられている。
東山哲也(ひがしやま・てつや)
1999年東京大学大学院理学系研究科修了(理学博士)。同大大学院理学系研究科助手を経て、2007年より名古屋大学大学院理学研究科教授。ERATO東山ライブホロニクスプロジェクトを主催、植物の受精システムを生きたままの細胞を捉える手法から研究している。














