RESEARCH
有性と無性を組み合わせて
多様性を維持するシダ
生きものは、遺伝的に多様な子孫を多く残すことで環境変化を乗り越え、続いていきます。通常、多様性を生みだすには有性生殖、素早く子孫の数を増やすには無性生殖が有利とされます。キナバル山のマレーホウビシダは、有性・無性生殖をうまく組み合わせた柔軟な生き方をしています。
1.有性生殖と無性生殖
私たち人間を含む多くの生物はオスとメスという"性"があり、有性生殖で次世代を残すが、"性"をもたない生物も少なくない。これを無性生殖とよび、子供はその親と遺伝的に同一のクローンとなる。この場合、集団内に他より少しでも適応度の高いクローンが出現すると、それが集団内に急速に広がり、最終的に集団内のすべての個体が遺伝的に同一のクローンとなる。そのため無性生殖種の遺伝的多様性は有性生殖種のそれと比べて低く、劇的な環境変化に対応できずに絶滅しやすいとされる。そこで無性生殖種を、いずれ滅びるものという意味をこめて「進化の袋小路」にはいった種とよぶこともある。しかし無性生殖は必ずしもデメリットばかりではない。有性生殖では交雑相手との出会いに多大な労力を要するが、無性生殖ではその必要がない。短期的には無性生殖種は有性生殖種と比べて多くの子孫を残す可能性が高いといえる。
2.シダ植物の無配生殖
シダ植物は通常、有性生殖をするが、なかには無配生殖(註1)と呼ばれる特殊な無性生殖をするものが存在する。有性生殖型では、受精後に葉をつくり、その後減数分裂を経て1倍体の胞子をつくり、また次の受精につながる(図1a)。一方、無配生殖では、胞子は2倍体で受精せずそのまま個体に成長する(図1b)。胞子が2倍体なのは、減数分裂前に核が融合して一時期4倍体になっているためである。つまり、遺伝的に同一のクローンをつくっているのである。無配生殖はシダ植物に報告例が多く、分類学の研究が進んでいる日本産シダ植物では、13%もの種が無配生殖をしている。分子系統学的解析から、この無配生殖はそれぞれ独立に有性生殖から進化したと考えられる。
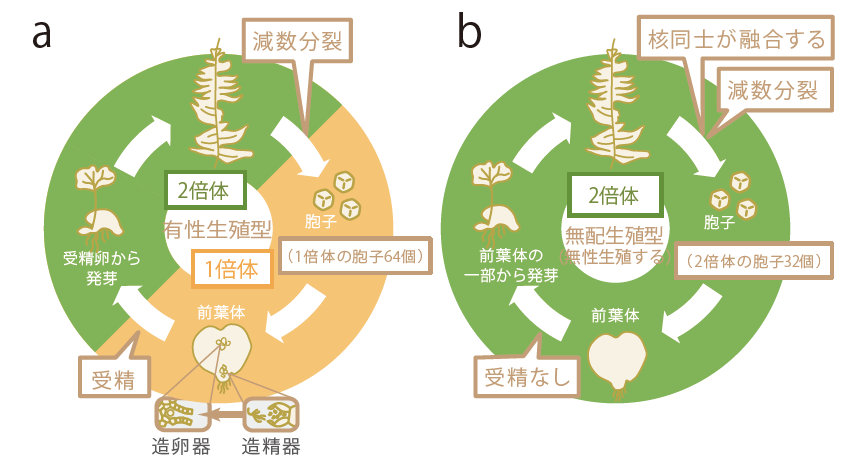
(図1)シダの生活環
有性生殖では、2倍体の植物体の葉の裏に胞子のうが形成され、1倍体の胞子が64個が生みだされる。胞子から前葉体ができ、前葉体上に造卵器または造精器がつくられ、卵と精子が受精し、2倍体の次世代個体が形成される(a)。それに対し無性生殖の一種である無配生殖は、体細胞分裂の途中で分裂しかけていた核同士が再度融合することで、2倍体の胞子が32個できる。その胞子からつくられた前葉体の細胞から直接次世代個体が発生する(b)。
(註1) 無配生殖
維管束植物において、配偶子が受精を介さず単為生殖によって単独に分裂・発達して胞子体を生じる現象。シダ植物の場合、胞子形成前に減数分裂を行うが、それより前に行われる体細胞分裂で核が融合する(復旧核形成)ため、生活環の中で倍数性が変化しない。
3.無配生殖型が遺伝的多様性を保つしくみ
近年、シダ植物の無配生殖型が詳細に調べられており、本来無性とされる無配生殖の生活環から外れた生殖様式をもつことがわかってきた(図2)。まず受精可能な精子をつくる能力があり、近くに近縁な有性生殖型の配偶体(前葉体)があれば交雑できる。二つ目に減数分裂時に染色体が不均等に分裂することである。無配生殖型の多くは3倍体であり、通常は3倍体の胞子体から3倍体の胞子をつくっているが、2倍体と1倍体の胞子を生じる不均等分裂をすることがある。しかも、ここで生じた胞子には(註2)稔性があり、次世代を残す能力がある。三つ目に、同一祖先に由来し潜在的に対合する能力をもっている染色体(同祖染色体という)同士が組み換えをする。3倍体の場合では3本の同祖染色体が存在し、これらの同祖染色体同士が減数分裂時に対合してしまう。この組換えで、親とは異なる新しい遺伝子型ができることになる。
ここに挙げた3つの現象は、いずれも実験室での人為的交雑で観察されたものであり、野外でこのしくみが実際にはたらいているかどうかはわかっていなかった。私たちがキナバル山で偶然発見したマレーホウビシダで実際にシダ植物の無配生殖種の遺伝的多様性を維持する姿が見えてきた。
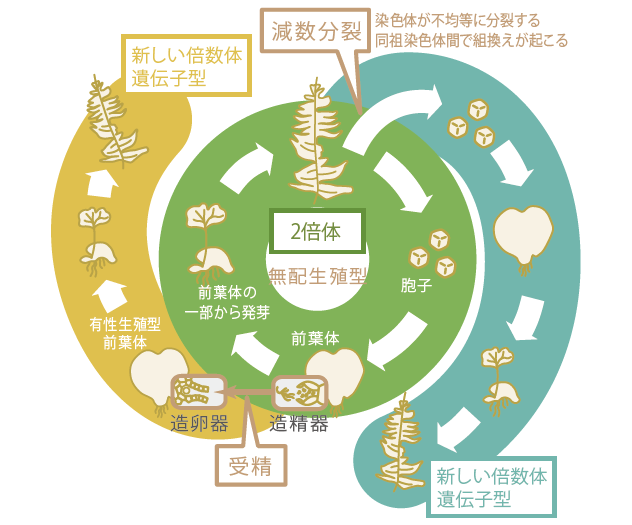
(図2) 多様性を保つしくみが見えてきたシダ植物の無配生殖
受精や不均等分裂、組み換えなどによって、無性的に増える通常の無配生殖の生活環から外れた暮らしをしていることがわかってきた。
(註2)稔性
有性生殖の過程で異常がなく、子孫を残すことができること。
4.キナバル山でみつかった謎のシダ植物
東南アジアで最も標高が高いキナバル山(標高4095m)には、裾野から中腹にかけて照葉樹が優先する自然林が広がっている(写真1)。2004年に中腹標高1500mから1900mの森の中で、私たちはこれまで報告例の無い形態形質をもつホウビシダ属のシダ植物を発見した。キナバルのシダ植物は詳しく調べられており、ホウビシダ属ではマレーホウビシダ、ウスイロホウビシダ、ウスバクジャク、ヤクシマホウビシダの4種が報告されていた。

(写真1) キナバル山
しかし、新しくみつけたホウビシダ属の種は、それらのいずれとも形態的特徴が異なっており、キナバル山では報告がないラハオシダという種に似ていた。ところが、DNAを調べたところ、ラハオシダではなく、マレーホウビシダに近縁であることがわかった。マレーホウビシダは東南アジアを中心に旧熱帯地域に広く分布しており、もちろんキナバル山にも生育しているが、新しいマレーホウビシダは非常に大きな葉をもっていた(写真2)。
同種の中で、このような形態変化がなぜ生じたのか? そこで、大型と標準型の葉を持つマレーホウビシダの細胞学的形質と遺伝学的形質を調べ、その理由を明らかにしようと考えた。

(写真2)標準型の葉のマレーホウビシダ(左)と大型の葉のマレーホウビシダ(右)。写真の中の定規は20cm。
5.種内で見られた多様性
マレーホウビシダでは有性生殖と無配生殖の2つの生殖様式が知られており、無配生殖型が広く分布していることがすでに報告されている。そこでまず、キナバル山のマレーホウビシダの生殖様式を調べた。有性生殖型は胞子嚢(のう)あたりの胞子数が64個であるのに対し、無配生殖型は32個となる。つまり胞子嚢(のう)あたりの胞子を数えることによって生殖様式が推定できるのだ。実体顕微鏡を現地に持ち込んで調べた結果、標準的なマレーホウビシダは無配生殖、葉の大きいマレーホウビシダには有性生殖と無配生殖の2種類の生殖様式が見られた。さらにそれぞれのタイプの染色体のセット数(倍数体)を調べたところ、標準型の葉で無配生殖をおこなう個体は染色体数117本で3倍体であり、大型の葉で無配生殖をする個体の中には155本で4倍体と117本で3倍体があり、有性生殖をする個体は78本で2倍体が見つかった(図3)。つまりキナバル山には形態と細胞学的形質で区別できるタイプが少なくとも4つあることがわかったのである。
これらのタイプの生育環境を比較したところ、標準型の葉で3倍体の無配生殖型は樹冠が開けた川沿いに、大型の葉で4倍体の無配生殖型は樹冠の閉じた林床の川沿いに、それぞれ生育していた。さらに大型の葉で3倍体の無配生殖型と2倍体の有性生殖型は川から離れた林道沿いの、やや乾燥した場所に同所的に生育していたが、詳しく見てみると3倍体は、より樹冠の開けた乾燥している場所に生育していることがわかった。4タイプ間では生育環境も異なるのである(図3)。
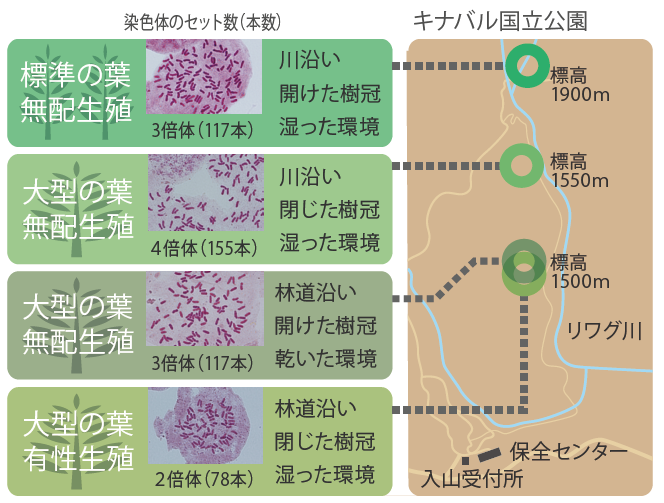
(図3)4つのタイプのマレーホウビシダ
赤く染まっているのは核内の染色体。大型の葉を持つ個体では、生殖様式や倍数性による違いが見られた。さらに生育環境も異なっており、全てで4つのタイプに分かれることがわかった。
4タイプの遺伝的関係を明らかにすべく、核由来の複数の遺伝子座と母性遺伝する葉緑体遺伝子の塩基配列を調べた結果、一部の大型の葉の3倍体無配生殖個体と4倍体無配生殖個体が、標準型の葉で3倍体無配生殖個体と大型の葉で2倍体有性生殖個体の雑種起源であることが明らかとなった(図4)。雑種である無配生殖個体の胞子は発芽し、前葉体を形成し次世代を残すことができる(写真3)。
また、葉緑体DNAの塩基配列から、この雑種の母親は標準型の葉をもつ3倍体無配生殖個体である可能性も示された。
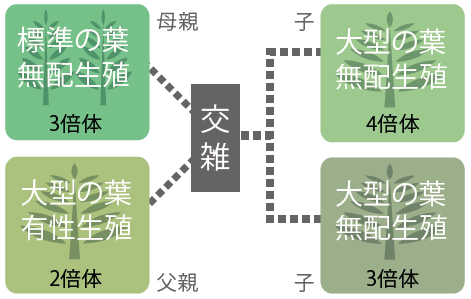
(図4)4つのタイプのマレーホウビシダの遺伝的関係
無配生殖型のマレーホウビシダには受精可能な精子の他に卵も備わっていれば、有性生殖型との交雑の他に無配生殖個体間の交雑も起こり、遺伝的多様性を集団に持ち込むことも可能となる。通常、交雑によって新しくできた遺伝子型は遺伝的浮動(註3)などにより、一定の確率で集団から取り除かれてしまう。しかし、マレーホウビシダの無配生殖型では、稀な交雑によって生まれた新しい遺伝子型もそれらの個体が無配生殖をすれば維持される。そのために無配生殖型が交雑と無性的な生殖を繰り返せば、有性生殖型よりも多型が維持されやすいと考えられる。
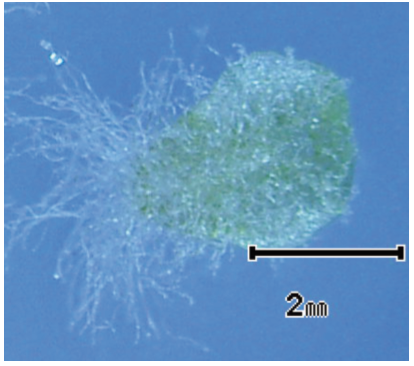
(写真3)雑種起源の無配生殖個体から生じた正常な前葉体。
(註3)遺伝的浮動
集団の遺伝子頻度が世代間で偶然的に変動すること。
6.無性生殖と有性生殖の組み合わせが生む多様性
無配生殖は無性生殖の一種であり、シダ植物でもそうであると考えられてきた。しかし私たちは今、シダ植物の無配生殖は、これまで考えられていたような単純な無性生殖ではなく、有性生殖等の遺伝的多型を生みだす能力と、大量に子孫を残せる無性生殖の能力の、両方を兼ね備えた生殖様式なのではないかと考えている。
外部形態からシダは1万種とされているが、遺伝的変異を獲得することで新たな環境に適応した無配生殖型や、交雑によって新たに生じた倍数体を含めると、その種類ははるかに多いかもしれない。今後、無配生殖型マレーホウビシダの自然集団で有性生殖能以外の遺伝的多型を生じるしくみ(減数分裂時の不均等分裂や同祖染色体対合)などが機能しているかを調べるとともに、他の分類群における無配生殖の多型を生みだす機構についても解析をすすめ、シダ植物の無配生殖という生殖様式の全貌を知り、種の多様性と生殖様式の進化との関係を明らかにしていきたい。
篠原 渉(しのはら・わたる)
2004年京都大学大学院理学研究科博士課程修了。理学博士。京都大学大学院理学研究科グローバルCOE特別講座助教。シダ植物と屋久島の高山性ミニチュア植物を対象に、種多様性、種分化、適応進化を研究している。














