RESEARCH
熱帯林樹木の多様性が続くしくみ
熱帯林に多様な生きものがいるのは当然と思われていますが、多様性は何によって決まり、どうやって続いていくのでしょう。マレーシアのキナバル山は、標高と地質の多様性に応じて、多様な森林が成立しており、山全体の植物多様性は世界最高レベルにあります。時間をかけてキナバル山の熱帯林を調査することで、熱帯林の骨組みを作る多様な樹木が共存するしくみが明らかになってきました。
1.多種が共存できる不思議
森林は、珊瑚礁とならんで、自然界でもっとも複雑な立体構造をもち、生物多様性が高い生態系と言ってよいだろう。その骨組みを作っている樹木は、光・水・土壌栄養塩(ミネラル)といった同じ資源を利用しながら、多種が共存している。これは一見何でもないように見えるが、実は不思議なことだ。
一般に、同じ資源をめぐって競争する生物種は共存できないとされ、例えばゾウリムシのような生物を使った飼育実験では、同じ資源をめぐって2種が競争すると、ふつう1種だけが生き残り、もう1種は絶滅してしまい、これは「競争排除法則」と呼ばれる。森林の樹木は、この法則に反しているように思える。樹木の多様性はどのようにして維持されているのだろうか? 世界でもっとも生物多様性が高い地域の一つ、マレーシアのキナバル山の熱帯林を例に、そのしくみを考えてみたい。
2.キナバル山の自然が生み出す多様な森林
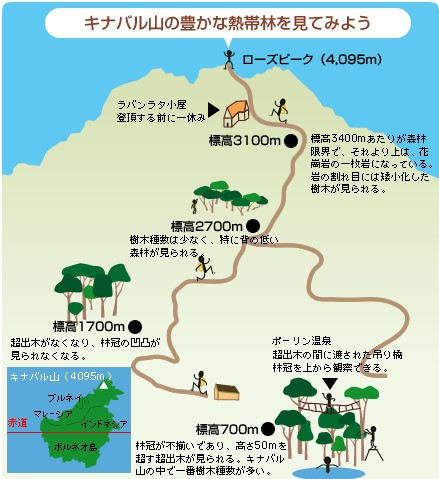
世界で三番目に大きい島、マレーシアのボルネオ島にあるキナバル山は、標高4095メートル、東南アジアで一番高い山である(図1)。島の海岸近くの低地では平均気温が27度もあり、典型的な熱帯気候となっている。しかし、キナバル山の平均気温は標高100メートルごとに約0.6度低下し、山頂では4度にまで下がり、これは北海道の平均気温に相当する。ただし日本と異なり冬がない熱帯では、落葉広葉樹林は存在せず、標高に応じた気温の低下とともに常緑の森林が単純化していくだけである。
キナバル山は、様々な地質が存在する点でもユニークである。標高約3000メートル以上の山頂部は花崗岩(註1)からなり、それより下の大部分は堆積岩(註2)からなるが、蛇紋岩(註3)も分布する。蛇紋岩は樹木の成長を抑制する特殊な化学組成をもつので、その上には小さい樹木が密生した特徴的な森林ができる。
以上のように、キナバル山は、高低差が大きく、さまざまな種類の地質によって形成されているため、標高と地質の多様性に対応して、多様な森林が成立し、多様性の高い植物相が育まれてきた。キナバル山周辺の1200平方キロメートルには、約5000種の維管束植物(シダ植物と種子植物)が生育する。日本全体とほぼ同じ種数が、300分の1の面積に詰め込まれていることになる。

(図1) ボルネオ島最高峰のキナバル山 (標高1700mの光景)
ボルネオ島のマレーシア領内にあるキナバル自然公園に、キナバル山がある。
(註1) 花崗岩
マグマが冷え固まってできた岩石の一つ。保水力が少なく栄養分は乏しいが、ミネラルの組成は第三紀堆積岩に似ている。
(註2) 堆積岩
泥や砂、生物の遺骸が積もり固まってできた岩石。
(註3) 蛇紋岩
岩の表面に蛇のような模様が見られることから命名された。マグネシウムを多量に含むため、植物の水分吸収能力を低下させる作用がある。
3.樹木の多様性を左右するもの
標高と地質の多様性に対応して植物の多様性が高いということは、場所によって樹木の種組成が大きく異なるということを意味する。私たちはまず、キナバル山で標高と地質の異なる9つの固定調査区(図2)を設定し、そこに存在する樹木をすべて同定し、共通種の割合に基づいて分類した(図3)。
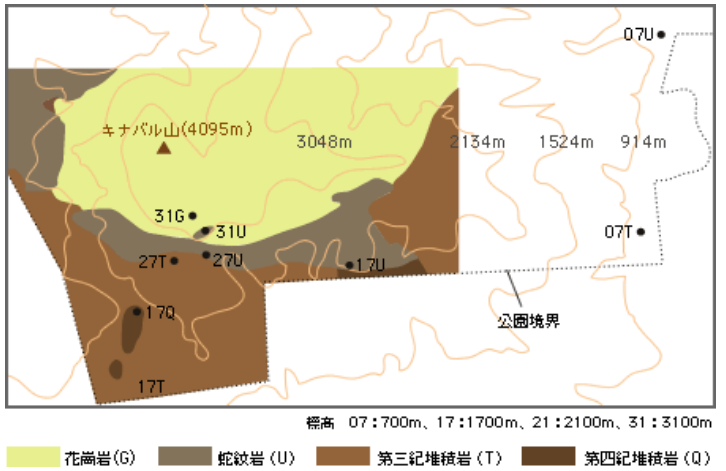
(図2) キナバル山に設定した9つの固定調査区
●は調査地点を、数字とアルファベットはそれぞれの標高と地質を表す。
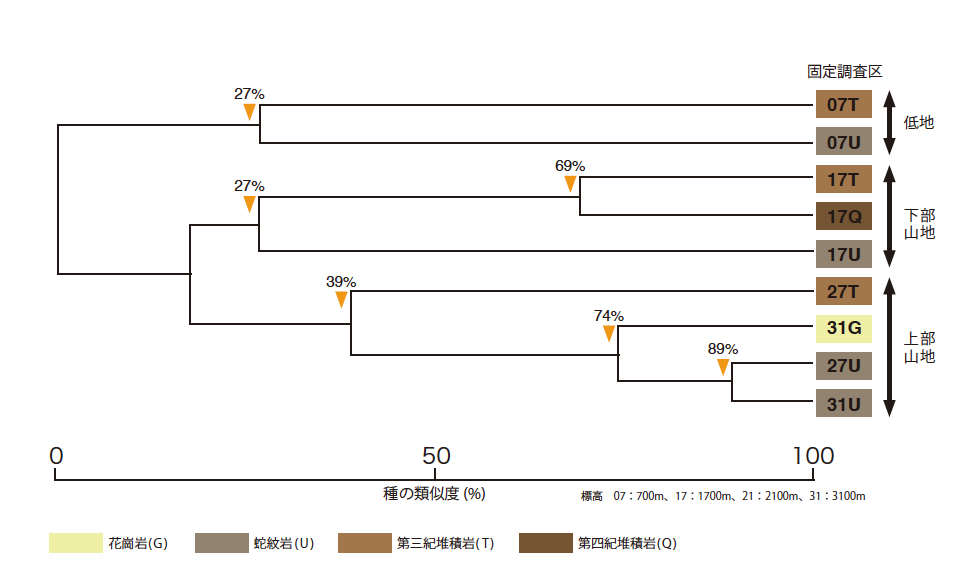
(図3) 標高と地質の違いによる種の類似度
固定調査区を共通種の割合に基づいて分類した図。横軸は、構成種の類似度を表す。
例えば、標高700mの堆積岩と蛇紋岩(07T, 07U)では、27%の類似度を示す。
調査区に存在するすべての樹木(幹直径が10cm以上)を構成種とし、その中で占有している幹の面積が大きい樹木を優占種とした。構成種に着目すると、それらは調査区の存在する標高の違いで大きく3つ(約700mの低地、約1700mの下部山地、約2700-3100mの上部山地)に分けられることがわかった(図3)。同じ標高で地質が違う場所を比べると、標高が高いほど構成種の類似度は高い。また、上部山地では、標高のわずかな違いよりも地質の影響のほうが大きくなる。例えば、標高2700mの蛇紋岩上の森林(27U)は、2700mの堆積岩上の森林(27S)よりも3100mの蛇紋岩上の森林(31U)に近い。
一方、優占種に着目すると、標高が高いほど類似度が低くなっていた。例えば、標高700mでは堆積岩(T)でも蛇紋岩(U)でもフタバガキ科が優占するが、標高3100mでは、花崗岩(G)でマキ科の針葉樹が優占するのに対し、蛇紋岩(U)では標高3300m以上の山頂部に多いフトモモ科の広葉樹が優占する。
生態系の多様性を扱う生態学では、一つの森林(調査区)に何種があるかをアルファ多様性、森林間でどれだけ種が異なるかをベータ多様性、この両者によって決まる地域全体の多様性をガンマ多様性という。今回の調査で、キナバル山でのアルファ多様性は熱帯林として必ずしも高くはないが、標高と地質の多様さがベータ多様性の高さにつながり、結果として地域全体のガンマ多様性は世界最高レベルにあるということがわかった。
4.森林の階層構造
キナバル山の森林構造を知るために調査区ごとに樹木の大きさを調査すると、種数の多い森林ほど樹木の背が高いことがわかった(図4)。
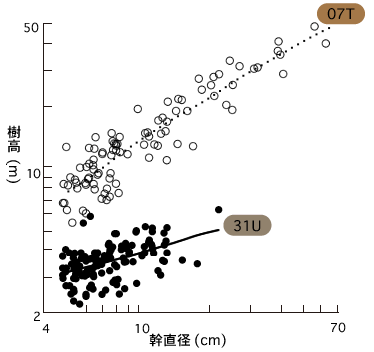
(図4) 調査区ごとに成育する樹木の樹高と幹直径
キナバル山で一番、樹木種数の多い700m堆積岩上の森林(07T)は、樹高が高く、幹直径も太い傾向にある。逆に、樹木種数が少ない3100m蛇紋岩上の森林(31U)は、樹高が低く、幹直径も細い。
種数のもっとも多い低地林では、周囲から突き出た樹冠を持つ種(フタバガキ科など)が存在するため、林冠(註4)が不揃いになる。これは超出木と呼ばれ、成長すると直径2メートル、高さ50メートル以上にもなる(写真1)。その根元には巨大な樹体を支えるための板根が発達し、周辺には数mの低い木が見られる。つまり低地林の森林は、超出木・林冠・下層という階層構造をもっているのだ(図5)。

(写真1)
幹直径2m、樹高50m以上にも達する超出木。
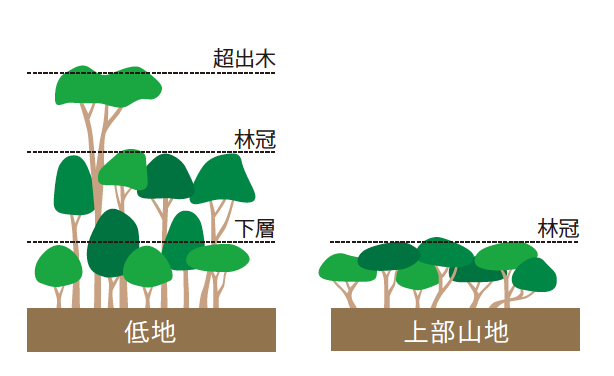
(図5) 森林の垂直構造
低地では、超出木、林冠、下層の3つの階層が生じ、高木と低木が共存できる。一方、上部山地では、林冠だけの一層となり、多種が共存できない。
低地林の次に種数が多い下部山地林では、超出木は見られず、外から見ると林冠の凹凸が少なく、日本の照葉樹林に似ている。樹高は30メートルほどで、板根のある木は少ない。種数がもっとも少ない上部山地林になると、樹高は10メートル以下にまで低下し、直径も30センチメートル以下のものが多くなる(写真2)。
つまり、低地では超出木・林冠・下層の3つの階層が見られるが、下部山地では超出木がなくなって2層になり、上部山地では林冠だけの1層になる(図5)。ここから、森林の垂直構造の発達が、複雑な階層構造をつくり出し、樹木の多様性を高めていると考えられる。

(写真2)
標高2700m蛇紋岩上の森林。樹高が低く、幹も細い。
(註4) 林冠
森全体を見た場合、一番上の面に枝と葉が集中している層。
5.森林の動きで生まれる多様性
樹木は動物のように自分で動くことはないし、その寿命は人間よりはるかに長いため、私たちは森林を静的なものとして考えがちである。しかし、継続して調査していると、森林もダイナミックに動いていることがわかる。
寿命が長いとはいえ樹木もいつかは死ぬ。また、突風で幹が折れたり、根こそぎ倒れることもある。このようにして森林に穴(ギャップ)ができると、そこで遷移(註5)が起こる。まず明るい場所ですばやく成長する陽樹が侵入し、ギャップを埋める(写真3)。ギャップが埋まると林床が暗くなるため、陽樹はそれ以上侵入できず、暗い場所でゆっくり成長する陰樹が育ち始める。やがて陽樹が枯れると、陰樹によって階層構造をもつ森林ができあがる。遷移は数100年かかって進行し、その過程で樹木の世代交代が起こる。実際には、このようなギャップ単位の遷移が、森林のあちこちでバラバラに起きているので、森林を上から見ると、様々な遷移段階の部分がモザイク状に組み合わさっているように見えるのである。ギャップ動態に起因する森林構造の変異を、水平構造の変異と呼ぶ。キナバル山でも、森林の水平構造は樹木の多様性を維持する重要な要素となっている。

(写真3)
太陽光がたくさん差し込むギャップでは、陽樹が優先的に育つ。
(提供:鈴木英治/鹿児島大学理学部)
(註5) 遷移
植物群落が時間の経過に伴って変化していく現象のこと。最終的には、安定な極相林となり、それ以上の遷移は進行しない。
6.多様性と生産性
ここで、多様な生態系は豊かな自然であるとはどういうことかを改めて検証してみよう。樹木は光合成をおこない、太陽の光エネルギーを固定し、二酸化炭素から有機物を作り出している。有機物は、樹木の幹や葉、あるいは花実となり、それらに依存する動物のエネルギー源となる。つまり、樹木の生産量が高ければ、より多くの動物を支えることができる。そこで、固定調査区内の樹木の幹直径を何年かおきに測定し、得られた増加量と落ち葉や花実の量(写真4)とを合わせることで、樹木の生産する有機物量(一次生産量)を推定し、さらに直径成長量と直径頻度分布から、樹木の世代交代の速さも推定した。

(写真4)
樹木の生産する有機物量を測定するための道具「リタートラップ」。この網によって、木の実や落ち葉などを回収する。樹木の幹には成長を正確に測定するため印がつけてある。(3100m蛇紋岩上にて)
キナバル山でこれまで15年にわたって蓄積したデータを解析した結果、樹木の多様性が高い場所のほうが生産性が高く、樹木の世代交代も速いことがわかった(図6)。前述の階層構造と合わせて考えると、生産性が高い森林ほど、垂直構造が発達して高木と低木の共存が促進され(図5)、水平構造の変動による陽樹と陰樹の共存も促進されるということになる(図7)。
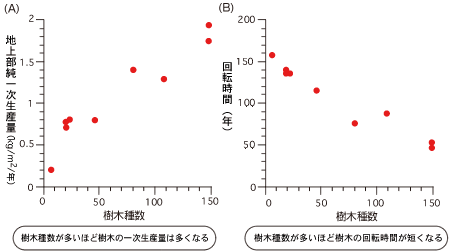
(図6) 固定調査区での樹木の生産性および回転時間
横軸は、1haあたり幹直径10cm以上の樹木種数を表す。(A)の縦軸(純一次生産量)は、光合成によって地上部に固定されたすべての有機物量を表す。(B)の縦軸(回転時間)は、幹直径10cm以上の樹木の平均余命を表す。
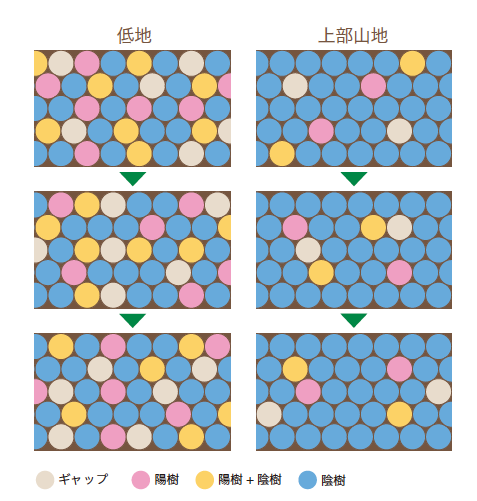
(図7) 森林の水平構造の変異
低地では、さまざまな遷移段階の部分が組み合わさり、陽樹と陰樹からなる多様な森林となる。一方、上部山地では、樹木の世代交代に時間がかかるためギャップが生じにくく、ほとんどが陰樹からなる森林となる。
7.樹木の多様性は必然と偶然によって続く
樹木の多様性はどのようにして維持されているのかという最初の問いに対し、キナバル山の熱帯林での研究から、それは森林の垂直構造と水平構造の変異によって動的に維持されていると答えることができる。特定の環境条件では、特定の種が有利だが、森林には空間的にも時間的にも様々な条件が作り出されている。つまり、森林は決して一様な条件にあるのではないので、たくさんの種が共存できるのだ。
ところで、最初の問いは一つの固定調査区での森林の種の共存というアルファ多様性について出したものであった。キナバル山での調査から、そこで特に高いベータ多様性はアルファ多様性とどのような関係にあるのだろうという問いが生まれた。 アルファ多様性は必ずしも高いとはいえないキナバル山だが、低地林や下部山地林では100種以上の樹木が1か所に共存している。これらすべての種が最適条件下に生育しているとは思えず、その種の多くは、生育最適地からの種子の供給によって、そうでない場所に生育しているのではないかと思われる。実はベータ多様性によってアルファ多様性が維持されているのだ(図8)。
つまり、森林における樹木の多様性は、森林構造に対応した決定論的プロセスだけでなく、種子散布という偶然に左右される確率論的プロセスにも大きく影響されると考えられるのである。今後は、両者のプロセスの関わりを明らかにし、それぞれのプロセスが多種共存に果たす役割を明らかにしていく必要がある。キナバル山での観測データをさらに積み重ね、この課題に取り組んでいきたいと考えている。
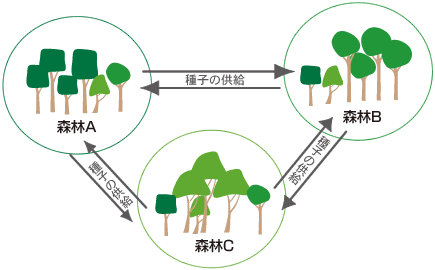
(図8) 種子供給によって維持されるアルファ多様性
森林ごとに繁茂している樹木が異なり、その種子が広く散布されることでα多様性が高まる。

相場慎一郎(あいば しんいちろう)
1997年北海道大学大学院地球環境科学研究科修了、博士(地球環境科学)。
日本学術振興会特別研究員、鹿児島大学助手を経て2007年より同助教。
.jpg)
.jpg)











