RESEARCH
ボルボックスで見る多細胞生物の形づくり
水中でくるくるまわる美しい緑藻ボルボックス。新しく誕生した個体は細胞のシートできた球状の体を反り返らせ、実に見事に裏返る。インバージョン(inversion:反転)と呼ばれるこの現象は、どのようなしくみで起こるのだろう。細胞のシートのさまざまな変化は動物の形づくりの基本だ。それがどのように行われるのか。この問いへの答を探るモデルとしてボルボックスは面白い。しかも多細胞化した時期が新しいので、単細胞生物から多細胞生物への進化の過程を見ることもできるのではないかと期待させる。
1.動物の形は細胞シートの変形から
動物の体づくりは、受精卵の分割から始まる。卵割と呼ばれる分裂を繰り返してできた球状の胚では隣り合った細胞がシート状につながり、その細胞シートが凹んだりたたまれたりして、原腸や神経管を経て複雑な器官が形づくられる。これは、ほとんどの動物の形づくりに見られるので、多細胞生物への進化の最も初期に現れた基本のしくみと考えられる。
このしくみを研究するモデルを探していた私は、思いがけず動物とは系統的に遠く離れた緑藻のボルボックスを用いることになった。その理由を説明するには、何はともあれ、ボルボックスとはどんな生きものかを知っていただかなければならない。
2.多細胞の球体ボルボックス(オオヒゲマワリ)
湖や沼に生息する緑藻の一つ。ボルボックス(Volvox)という名前がラテン語で「勢いよくころがるもの」を意味し、和名がオオヒゲマワリというところからも想像できるように、きれいな球形で回転しながら泳ぐ。(図1)。
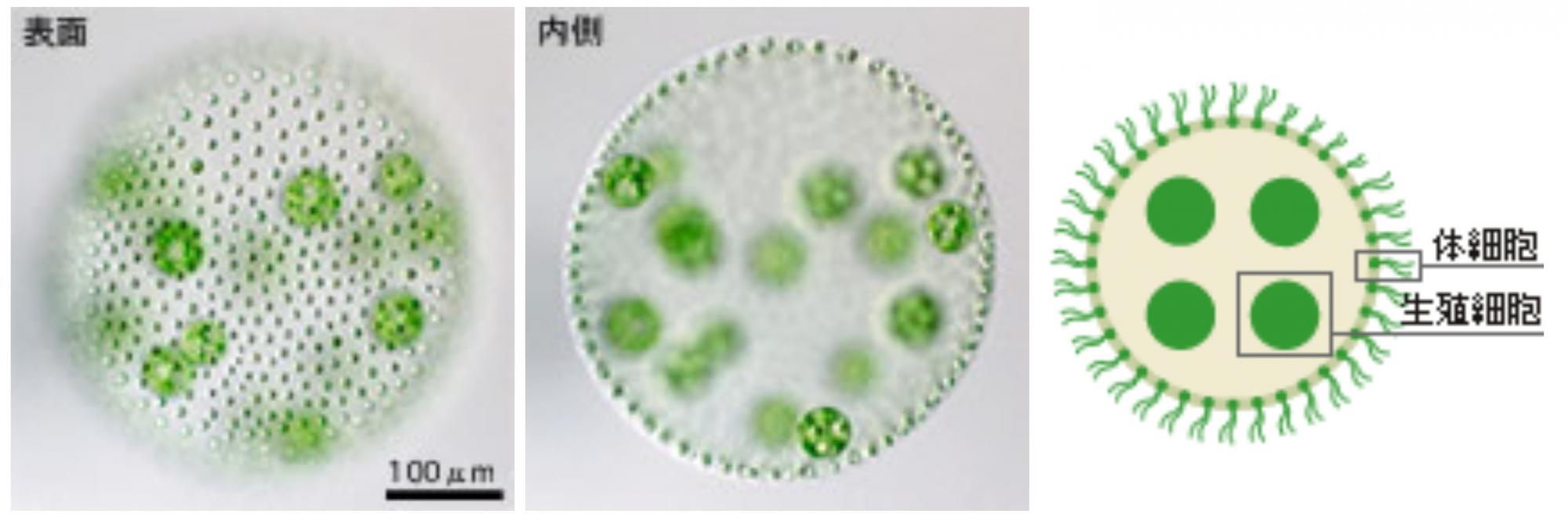
(図1) ボルボックスの体のつくり(成体)
実験に用いたボルボックス(V. carteri)は、直径0.5ミリの球体で、約2000個の小さな細胞(体細胞)が球体の表面に一層に並ぶ。その内側に16個の大きな細胞(生殖細胞)がある。体細胞の2本のべん毛の運動が統合され、水をかいて泳ぐ。
体のつくりは単純で、よく研究に用いられるV. carteriでは、約2000個の小さな細胞が球体の表面に一層に並び、その内側に16個の大きな細胞を包みこんでいる。小さな細胞はみな2本の鞭毛を生やし、その運動が統合され、回転して泳ぐ。この小さな細胞が2本の鞭毛をもつ単細胞生物クラミドモナスとそっくりなので、それが集まって生活していると勘違いされることが多い。群体と呼ばれることもそれを助長しているのだろう(*註1)。しかし、ボルボックスはれっきとした多細胞生物で、球体の内側の16個の細胞は生殖細胞だ。この生殖細胞が分裂し、親の球体の中で次の世代が生まれていくのだが、分裂の途中で不等分裂が起こり小さな細胞(体細胞)と大きな細胞(生殖細胞)が生み出される(図2)。このやり方は、動物の卵割とよく似ている。分裂を終えた子どものボルボックスはやがて形づくりを完成し、親の球体を破って水中に飛び出る。
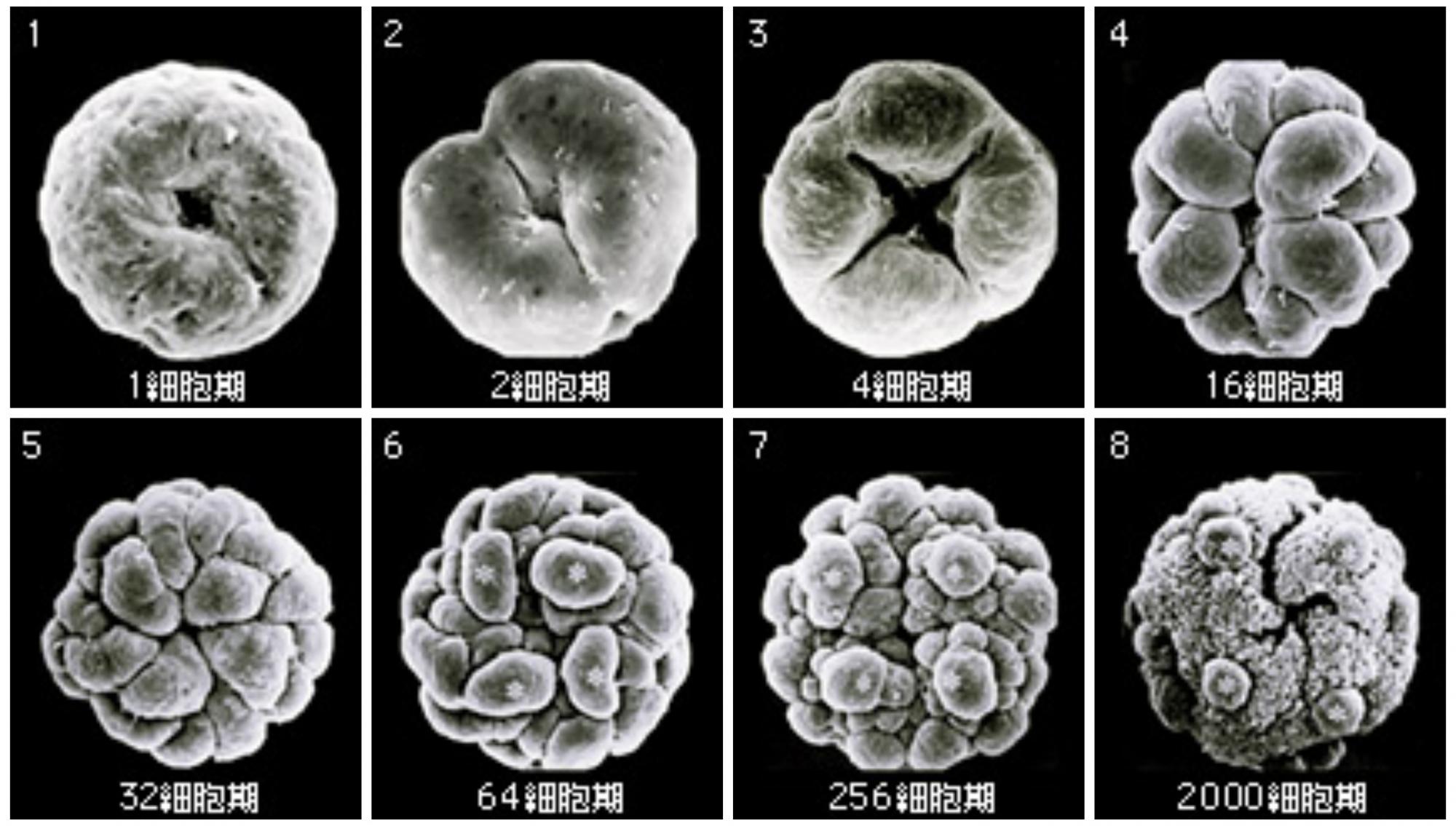
(図2) ボルボックス胚の分裂
分裂直前の生殖細胞から32細胞期までは等分裂で全ての細胞の大きさは同じ。64細胞期になると不等分裂で大きな細胞が生まれ(*で示す)、256細胞期では大きな細胞と他の細胞の差がより明確になってくる。分裂を完了した胚は成体と同じ約2000個の細胞を持つが、大きな細胞(生殖細胞)は外側にあり、成体とは表裏が逆転している。
(走査電子顕微鏡写真:D.L. Kirk博士 )
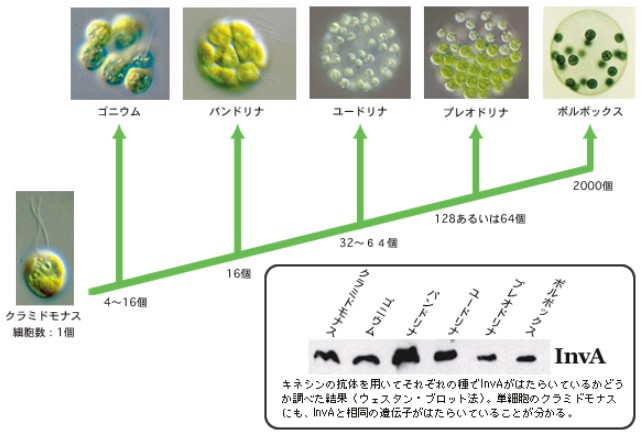
(註1)単細胞生物クラミドモナスから多細胞生物ボルボックスへの進化
DNA解析をもとにした分子系統樹から、ボルボックスはこの5000万年の間に、単細胞生物のクラミドモナスから進化したと推定されている。さらに、体を形づくる細胞の数と分化の程度が中間的なゴニウム、パンドリナ、ユードリナ、プレオドリナ等が存在する。
3.飛び出す前に裏返し
分裂を終えるまでのボルボックス(胚の段階 *図2の2000細胞期)と、親のボルボックス(成体の段階 *図1)とを比べると、ふしぎやふしぎ球体の表裏が逆になっている。成体で内側にあるはずの生殖細胞が、胚では外側にあるのだ。実は、胚から成体への変化の際に、くるりと裏返る、インバージョン(inversion:反転)と呼ばれる現象が起こるのだ。
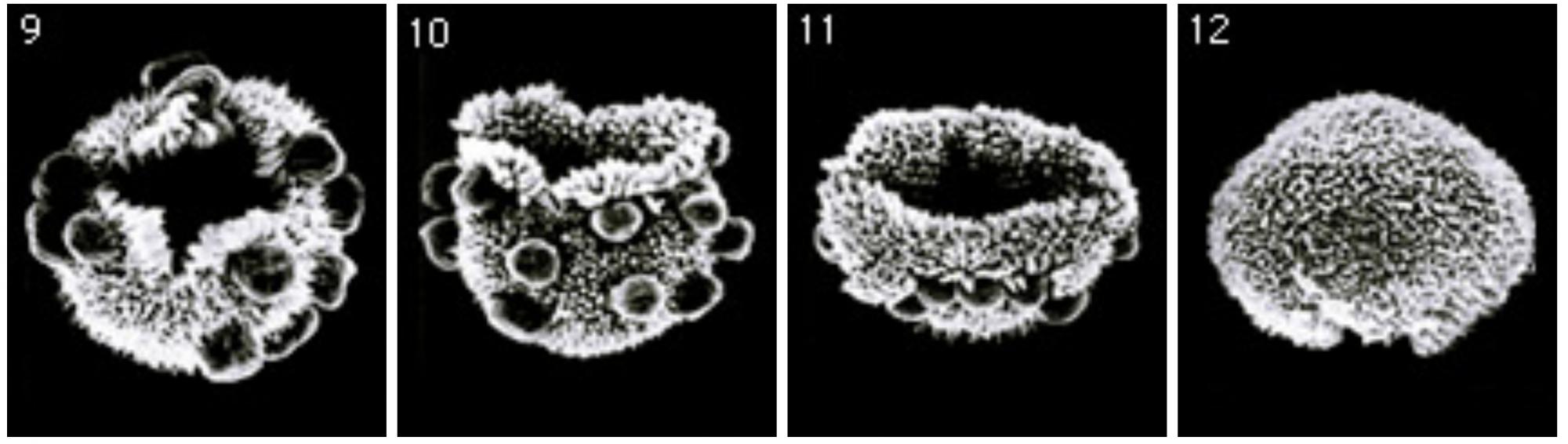
(図3)ボルボックス胚のインバージョン
分裂を完了した胚(図2-8)の一方の極には、十字に切れた穴があり、その穴の周りが外側に反り返ることから、インバージョンは始まる。反り返り運動がもう一方の極へと徐々に伝わり、最後には表裏が逆転すると、生殖細胞が内側へと入り、成体と表裏が同じに胚なる。(走査電子顕微鏡写真:D.L. Kirk博士 )
実際にインバージョンを見て頂こう(図3)。ボルボックスの胚の一方の極には、十字型に切れた穴(フィアロポアー)があり、その穴の周りが外側に反り返ることから、インバージョンは始まる。反り返り運動がもう一方の極へと徐々に伝わり、最後には表裏が逆転し、生殖細胞が内側へと入って手品は終り。
体細胞にとってもこの現象は重要だ。胚の段階では、体細胞の鞭毛は内側に向かって生えているが、インバージョンによって外側に向く。外に向かって伸びた鞭毛で水をかいて泳げるようになるわけだ。体細胞は接着性のタンパク質を分泌し、隣り合った細胞間を埋めることで成長し、やがて親の球を破って水中に飛び出す(図4)。
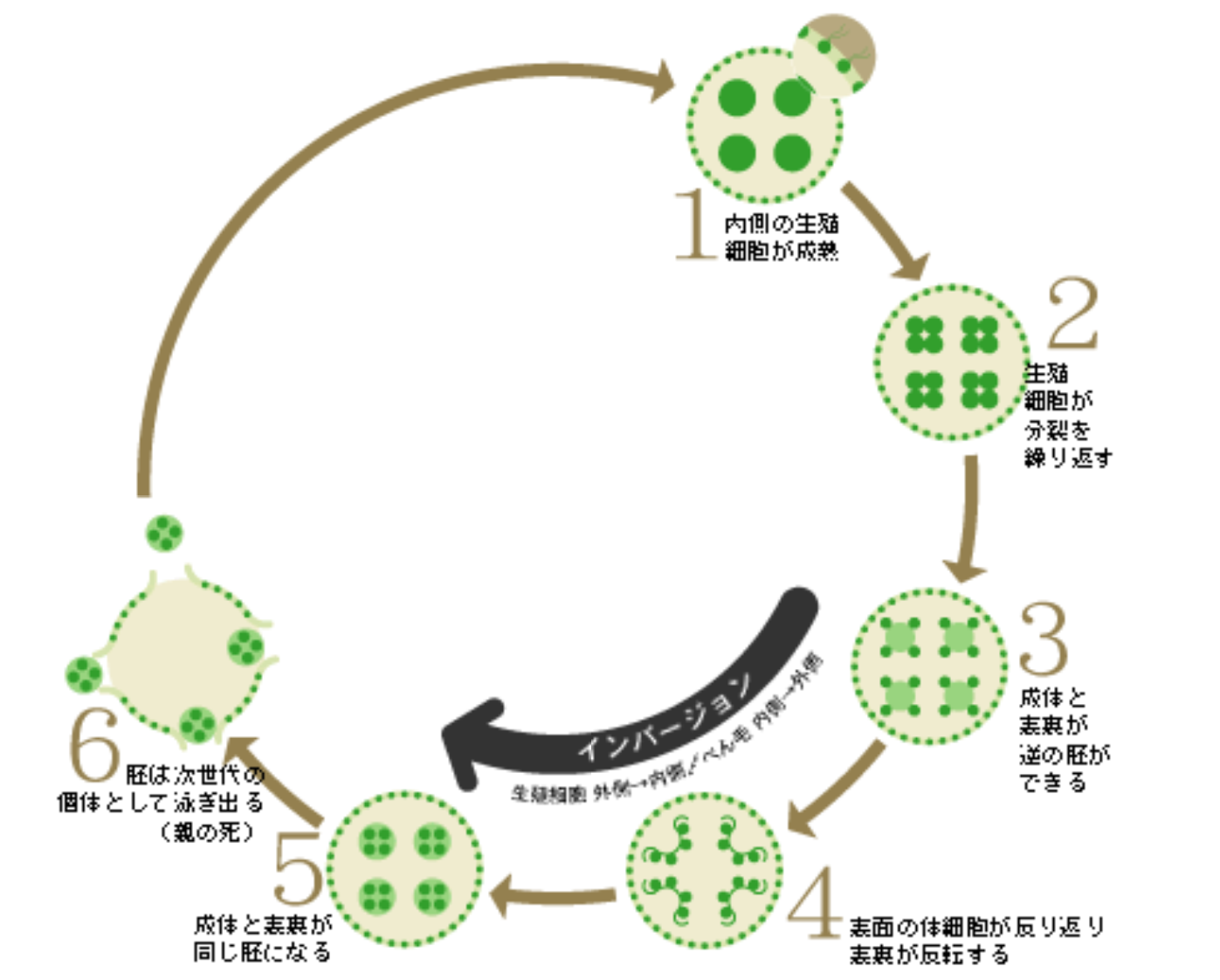
(図4) ボルボックスのライフサイクル
4.ボルボックスが形づくりのモデルになる理由
ボルボックス胚のインバージョンが、細胞シートの変形を研究するよいモデルになると考えた理由は二つある。まず、ボルボックス胚の構造は単純だ。十数個の生殖細胞を除けば、同じ種類の体細胞が一層に並んでいるだけで、細胞シートとして見た時、動物の胚と比べて非常に単純だ(基底膜などの細胞以外の構造が無い)。ほぼ究極の単純さと言ってよかろう。実はこれに加えて、ボルボックスならではの利点がある。インバージョンに異常をもつ突然変異体が容易に得られることである。細胞シートの変形は形づくりの最も基本的なしくみであるため、動物ではそこに関わる遺伝子が欠損すると生存すること自体が難しい。しかしボルボックスでは、インバージョンより前に生殖細胞ができているので、インバージョンができなくても次世代を残すことができるわけだ。これでなぜボルボックスを研究材料に選んだかはわかっていただけたと思う。
私が師事するD.L. Kirk博士(米国Washigton大学・教授)のグループは、四半世紀ほど前にボルボックスのインバージョンの研究を精力的に進めた。まず、インバージョンを始めた細胞シートでの一つ一つの細胞の形態を走査型電子顕微鏡で観察した。反り返る前の細胞は紡錘形をしており、隣り合う細胞は中央で接着している(図5-1)。ところが反り返りが起きたところでは、細胞の一端が胚の外側に向かって伸びて瓶型に変形し、その柄の先端で隣の細胞と接着している(図5-2)。つまり、紡錘型から瓶型へと細胞が細長く伸びる形の変化と、隣り合う細胞をつなぐ接着部位(ブリッジあるいは原形質連絡と呼ぶ*図5矢印)の移動という、2つの段階が共役してはたらいていることがわかる。この移動は、中央にあったブリッジが切れて先端に新しいブリッジが生まれるのではなく、ブリッジに対して細胞がずれていくことで起きる。このようなはたらきが十字型の穴の部分から順番に起こることで細胞シートが徐々に反り返り、最終的に裏返しになるのだ(図6)。
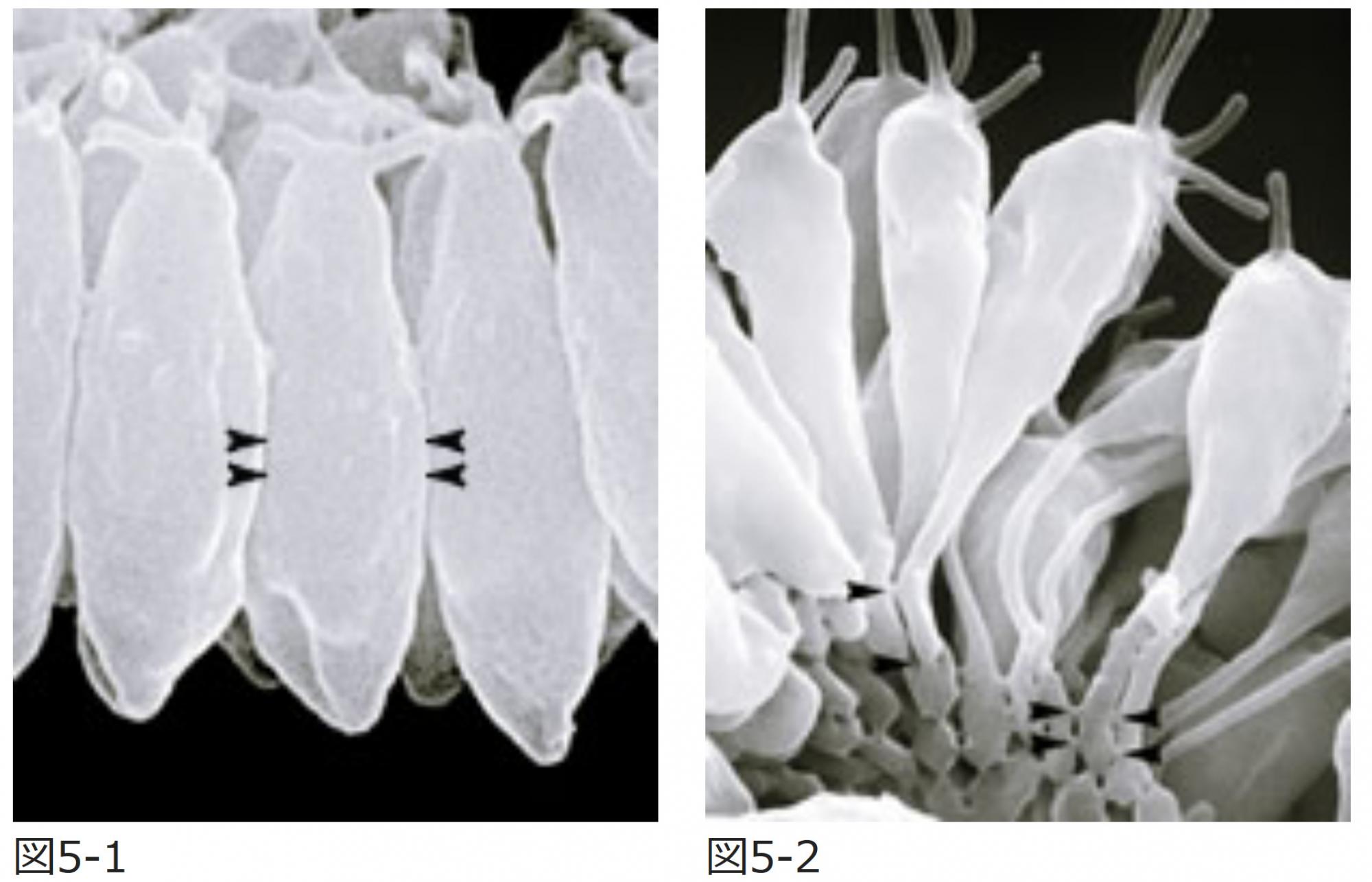
(図5) インバージョンを引き起こす細胞の変形とブリッジの移動
反り返る前後の細胞の電子顕微鏡写真。反り返る前の領域(図5-1)の細胞は紡錘型で、隣り合う細胞は中央でつながっている。一方、反り返りの起こっている領域(図5-2)の細胞は長い柄を持つ瓶型で、隣り合う細胞は瓶型の柄の先端でつながっている。矢印 は、細胞間の接着部位(=ブリッジ/原形質連絡)の位置を示す。

(図6)インバージョンの運動モデル
インバージョンの始まった開口部では、細長く伸長する細胞の変型とともに、ブリッジ(モデルの赤い部分)に対して細胞がずれていく。その結果、細胞シートは折れ曲がる。このような細胞の変形とブリッジに対する細胞の移動が開口部から順番に起こればインバージョンをうまく説明できると考えられる。
5.インバージョンに関わる遺伝子
次なる作業は、突然変異体を用いて、このメカニズムに関わる遺伝子を追うことだ。最近、私の研究によって非常に興味深い遺伝子invAのはたらきを明らかにすることができたので、紹介しよう。インバージョンが途中で止まってしまう変異体(InvA)を観察すると、細胞の形は紡錘型から瓶型に変化しているが、細胞をつなぐブリッジの移動が起きていないことがわかった(図7)。

(図7) InvA変異体
InvA変異体は、インバージョンが途中で止まる。図7は、反り返りの止まったInvA変異体の電子顕微鏡写真。図5の野生型と比較すると、細胞は瓶型に変形しているが、ブリッジの移動が起こっていない。
そこで、変異の起きている遺伝子invAを単離し、DNA配列を調べたところ、キネシン (*註2)というタンパク質をつくる遺伝子だった。さらに、ボルボックスの胚でこのキネシンがどこにあるか調べると、図6のブリッジの移動パターンと一致し(図8左)、反り返りの前後で拡大して見ても、紡錘型細胞では中央部に、瓶型細胞では柄の先端にと、キネシンはブリッジと共に移動することがわかった(図8右)。
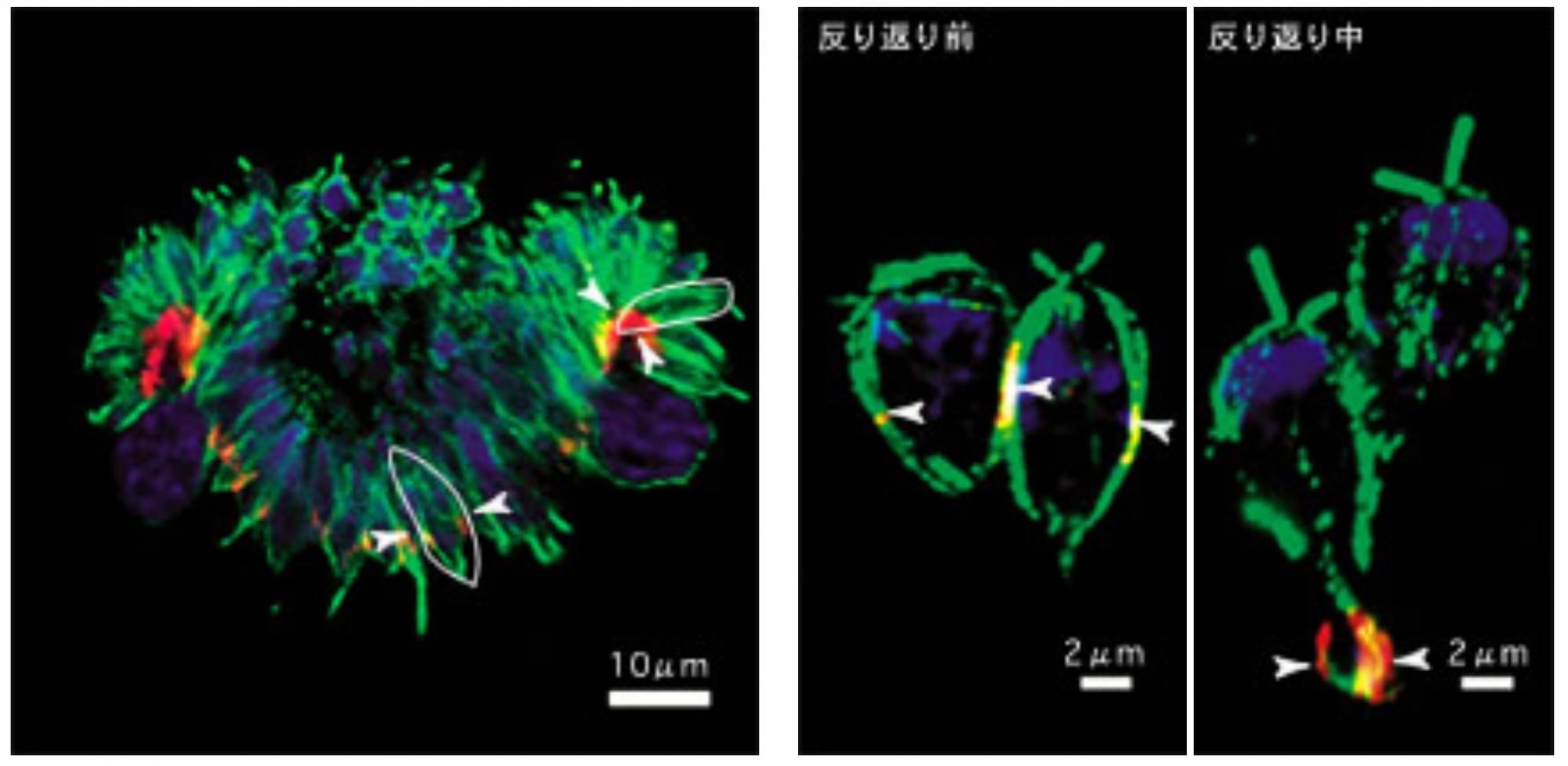
(図8)インバージョン胚でのキネシンの移動
抗体によって染色したインバージョン中の胚の顕微鏡写真。赤く染まった部分がキネシン(矢印で図示)。緑に染まった部分は微小管で、1つ1つの細胞の輪郭に一致する(白い囲みで図示)。青に染まった部分は核DNA。左図は胚の全体像を、右図は反り返りの前後で細胞1つ1つを拡大して見ている。
キネシンは微小管とよばれる繊維の上を一方向へ移動する分子モーターである(*註2)。細胞の形を支持する骨格の役目をもつ微小管は、ボルボックス胚の細胞では細胞膜を裏打ちし、ブリッジの近くにも走っている。微小管には極性があり、瓶型細胞の柄の先端がプラス極で、反対側(基底小体の方向)がマイナス極であることがわかっている。アミノ酸配列から、このキネシンはマイナス極からプラス極へ移動するモーター活性をもつと推測できた。つまり、ブリッジの移動する方向に一致する。このことから私は、遺伝子invAのはたらきを説明する次のようなモデルを考えている。インバージョンの全過程を通して、invAがつくるキネシンはブリッジに結合しており、ブリッジ近くの微小管と相互作用できる距離にある。キネシンがプラス極(柄の先端)へ移動しようとすると、相対的に微小管をマイナス極(基底小体)の方向に押し上げ、細胞をブリッジに対して滑らせる。この結果、ブリッジが柄の先端に移動し、細胞シートの反り返りが起こるというものだ(図9)。
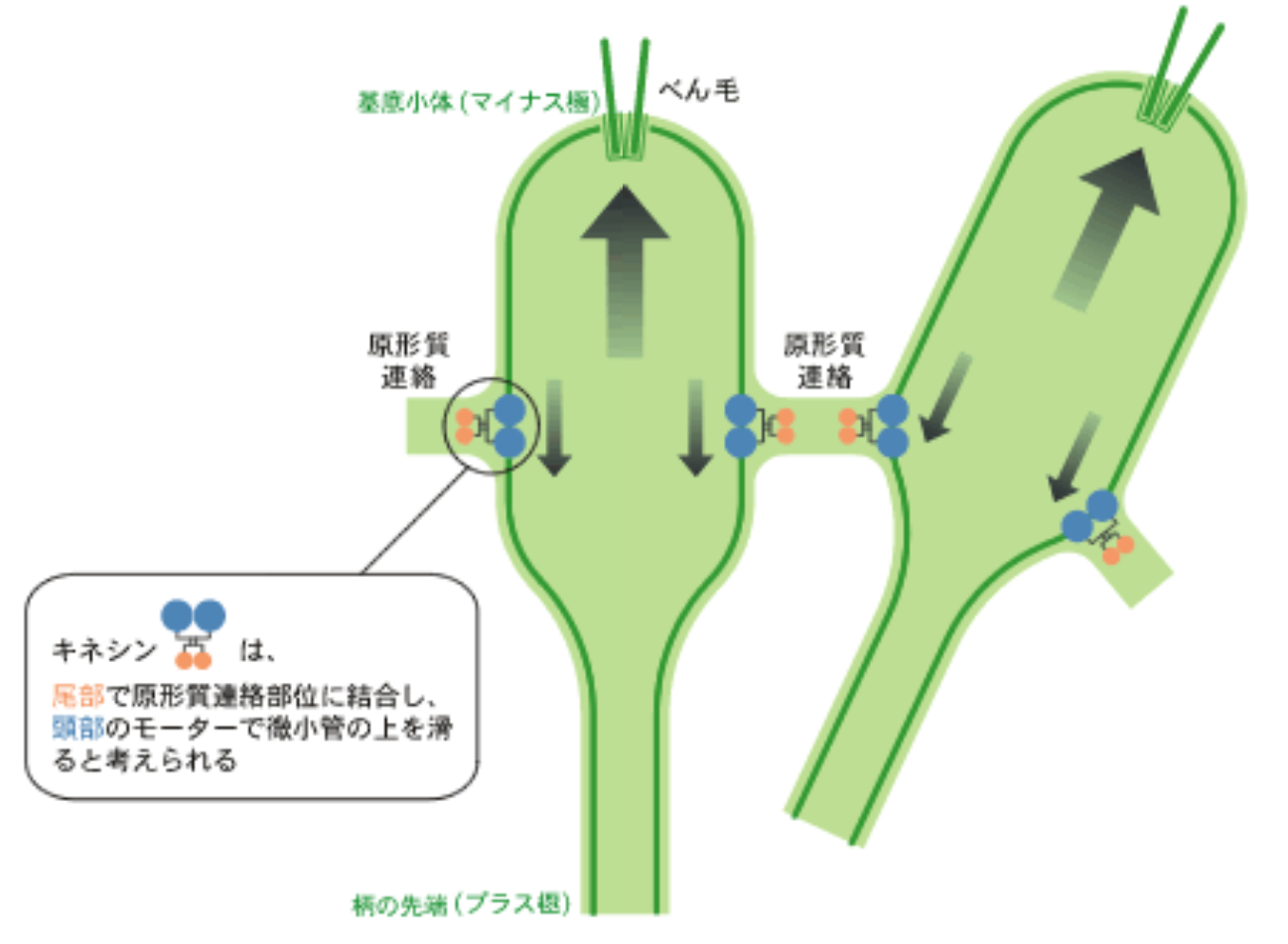
(図9)インバージョンの運動モデル
インバージョンの全過程を通して、遺伝子invAがつくるキネシンはブリッジに結合しており、ブリッジ近くの微小管と相互作用できる距離にある。キネシンがプラス極(柄の先端)へ移動しようとすると、相対的に微小管をマイナス極(基底小体)の方向に押し上げ、細胞をブリッジに対して滑らせる。この結果、ブリッジが柄の先端に移動し、細胞シートの反り返りが起こると考えられる。
(図1) キネシンと微小管
微小管は真核生物に共通して存在し、細胞の形を支持する骨格繊維である。分子モーターは微小管をレールとして使い、自身と結合したさまざまな物質を輸送する。このとき、微小管の繊維にはプラスとマイナスの極性が決まっており、分子モーターの一種であるキネシンの多くはプラス端に向かって移動する。
動物の脳をつくる神経細胞でも(※関連記事リンク:生命誌10号 Special Story「生命をささえる運び屋分子 廣川信隆」※)、軸索の中には微小管が何本も走っており、キネシンが神経伝達物質やシナプス小胞を輸送している。緑藻のボルボックスと動物の脳で、同じ骨格繊維と分子モーターがはたらいているとは。遺伝子の融通がきくのには驚く。
6.多細胞化への進化が見える?
DNA解析をもとにした分子系統樹から、ボルボックスは単細胞生物のクラミドモナスからこの5000万年の間に進化したことがわかっている。動物や陸上植物の多細胞化は約5億年以上前の先カンブリア時代とされているが、このように最近になって(5億年と比べれば!)の多細胞化もあるということは興味深い。多細胞化は決して進化の過程でただ一度起きただけのことではないのだ。多細胞化からの時間が短いということは、中間的な形をもつさまざまな種が現存していることであり、進化の過程を追うのに適しているかもしれない(*註1-囲み部分)。
インバージョンする細胞シートの動きにとって重要なはたらきをするボルボックスの遺伝子invAと非常によく似た遺伝子が、インバージョンを行わない単細胞のクラミドモナスでもはたらいていることが分かってきたのも面白い。脳ではたらく遺伝子がカイメンにすでにあるなど、遺伝子とはなにかという本質的な問いがさまざまな研究から出ている今、これがなにを意味するのか、じっくりと考えようと思っている。
進化とは過去の出来事であり研究の対象としては難しい。分子生物学や細胞生物学といった学問に慣らされてきた私は、少し前までは、そのように考えていた。しかし、ボルボックスの研究を続けているうちに、単細胞生物から多細胞生物が誕生する進化の時がビデオの早送りのように浮き彫りになるのではないかという期待がふつふつと沸いてくる。形づくりの基本は何で、いつ始まり、何がきっかけで、どんな風に変わってきたのか。さあ見せてくれよと、フラスコの中のボルボックスに語りかける日々だ。
西井一郎(にしい・いちろう)
1999年大阪大学大学院理学研究科生理学専攻博士課程修了。1999年より日本学術振興会海外特別研究員として、米国セントルイスWashington大学のDavid Kirk教授の下にてポストドクトラルフェロー。自作の「反り返って裏返る球体」模型を携え、ボルボックスの研究を広めようと行脚中。














