Lecture
進化のポリフォニー
東京都立大学の石川良輔教授(理学部自然史講座)は、三十数年にわたって日本全国のオサムシを採集し、詳細な分布マップを作ってきた。交尾器の変化というこれまで見過ごされていた進化の指標に注目し、従来の分類体系を大幅に塗り替える新しいオサムシの進化の道すじを描き出した。石川教授は、日本におけるオサムシの進化を箱庭的進化と呼ぶ。私たちはこの小さな日本のおかげで、進化という生き物のダイナミックな時間的変化を、縮図的なこまやかさで見ることができるのである。
宝石の輝き

1-1(左)、1-2(右)世界のオサムシ
ヨーロッパを左、アジアを右にして、ほぼ世界地図の北半球に合わせて、世界のオサムシを並べてみた。
1-1(左)中ほどにある玉虫色の美しい一群は、中央ヨーロッパからフランスに分布するクリソカラブスの仲間。その右手一番下のやや地味な個体はイタリア・ピエモンテの高地にしかいない希少種。そのすぐ右上の美しい個体は、戦乱のさなかにあるセルビア産。中ほどの大きな個体二匹は、オサムシ群中最大のフトオサムシで、上の緑色がコーカサス、下の紫色がトルコ産。
1−2(右)中央部から上は、中国からシベリアのツンドラ、タイガ、ステップに分布している種。ヨーロッパのもののようには目立たないが、希少種もあり、土地柄を示す。上列左隅の美しい個体は西シベリア・セミパラチンスク産。原爆試験地でいまでは近づけない。下側には、中国で著しく分化したカブリモドキの一群と、トゲオサムシ(最下列左から4、5)が並ぶ。カブリモドキの背のこぶこぶが印象的。その上は朝鮮半島のカブリモドキ。右端の下から二番目は日本の固有種マイマイカブリ。右端上から三番目が、日本種では珍しく宝石色の輝きをもつオオルリオサムシ。左下方にパキスタン、カシミールあたりの種が並び、その上にイラン周辺の中央アジア産が並ぶ。
東京・原宿の石川教授宅を訪れると、オサムシの膨大な標本の数に驚かされる。30年間で訪れた採集地は1000ヵ所以上。採ったオサムシは5万匹にのぼる。大型のドイツ箱だけで90箱もあり、合わせて数百箱が部屋の中に所狭しと置かれている。一つの種類だけで1万匹近いものが何種類もあるという。
オサムシは、カブトムシやクワガタムシと同じ甲虫の仲間だが、目立った角や大あごがあるわけでもない。とくに日本産は色彩も一部を除けば地味である。虫の熱心な収集家は、アマチュアのマニアに劣らず生物学者の間にも多いが、蝶やカミキリムシなどに比べれば、オサムシは圧倒的に「玄人好み」(専門家の意味ではなく、収集家の中のさらに好事家の人たち)の昆虫といっていいだろう。

アオオサムシ(東京・狭山湖畔)
しかし、この何の変哲もないと思っていた虫が、標本箱に集められた無数の種類を見せられた途端に、ため息が出るほどの迫力で迫ってくる。色彩は決していわれるほど地味ではない。濃紺の深い青色から、濃い緑、さらには赤銅色へとさまざまな変異を見せる。北海道にいるオオルリオサムシは、まさに瑠璃色の輝きをしている。
目を海外のオサムシに転ずると、その美しさには呆然とするほかはない。中央ヨーロッパからフランスまで広く分布しているクリソカラブス・アウロニテンスの一箱を開いたときには、そのまばゆさに息を飲んだ。なんという青、なんという緑、なんという紫、としか表現できない色彩のパレード。
別の箱に収められているトルコのオサムシの一群は、日本のオサムシが2~3cmなのに比べ5~6cm近い大型である。色は地中海を思わせるような深い、深い青。ただ、見事としかいいようがない。ヨーロッパー帯では、オサムシは美しい甲虫として一般の人にもよく知られており、日本の蝶のようにアマチュアにもたいへん人気のある虫なのだという。
2. オサムシ
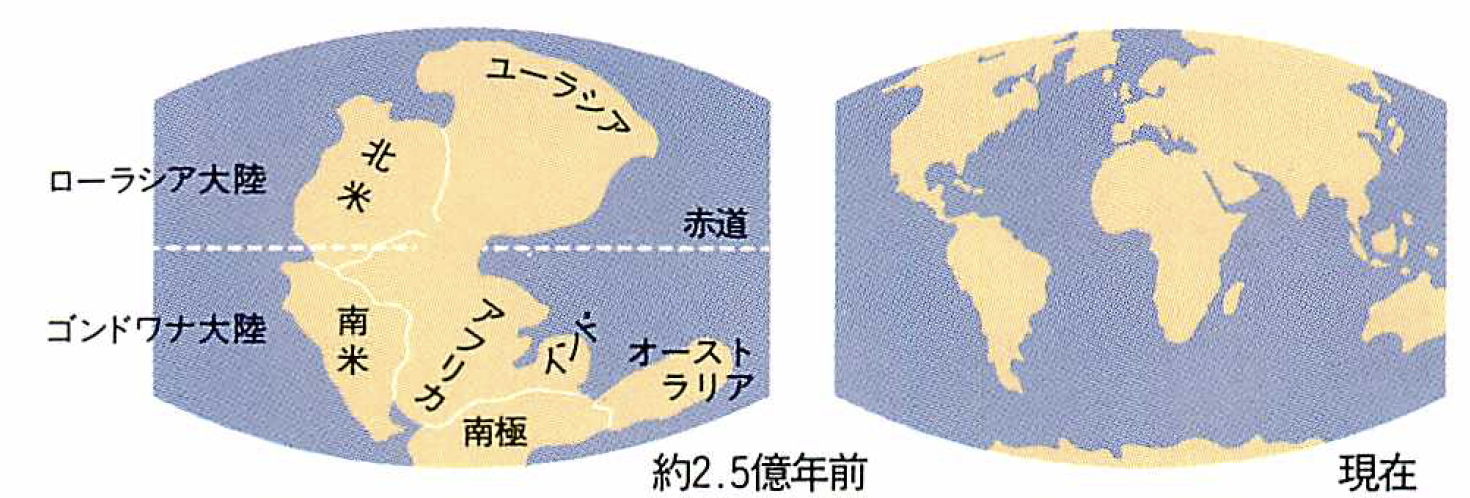
クワガタムシやカミキリムシと同じ甲虫の仲間。ヨーロッパ、インド南部を除くアジア、北アフリカ地中海側、メキシコ以北のアメリカに分布。世界で約700種が知られている。地質学的には、約2億年前の中生代に分かれた二つの大陸ゴンドワナとローラシア(地図参照)のうち、ローラシアの部分に分布域が一致している。ごく少数の例外を除いて飛ぶことができない。このため移動する範囲がかぎられているので、地理的な変異が生まれやすいと考えられている。
進化の道すじ
生物が時間の流れの中で、さまざまに分岐し多様化していったことは疑いないとしても、その多様化の道すじ、つまり「進化の道すじ」は必ずしも明確ではない。それは一つには、種の概念に暖昧性があり、種と種を区別する明確な指標に欠けていたからではないだろうか。オサムシにしても、種や亜種といった区別は大型とか小型、あるいは体表の彫刻差など、主に形態上の違いだけでなされてきた。こうした分類方法では、変異の時間的な方向性があまり見えてこない。その結果、なぜあるオサムシがそこにいて、別のオサムシがあそこにいるのかといった空間的な関係も不明のままとなる。
時間の流れの中で、一つの方向性を持って変化してきたような「指標」が見つかれば、それはたんに種の区別だけでなく、進化の道すじをも示していることになるだろう。石川教授は、交尾器の形の変化の系列から、少数のオサムシ祖先型が、日本の各地へと拡散し多様化していった道すじを、地図の上へと描き出したのだ。

一般的に山や川は、オサムシたちの地理的境界になることが多い。
30年以上前になるが、石川教授は奈良の金剛山で奇妙なオサムシを採った。外形は関西地方の平地でごくふつうに見かけるヤコンオサムシにそっくりなのに、雄のペニス(陰茎)が2倍以上も大きいのだ(図6参照)。その長さはじつに体長の半分にものぼる。
オサムシの雄のペニスには内袋があって、交尾するときにはこの内袋がペニスの先端から反転して突出し、雌の膣に入る。この内袋には、交尾片と呼ばれるキチン質の骨片がついていて、やはり雌の膣に入っていく。驚いたことに金剛山のこのオサムシでは、骨片がさらにペニスと同じくらい大きいのである。
石川教授は、このオサムシを新種と見て「ドウキョウオサムシ」と名づけた。もちろん、ドウキョウとは、巨大な男根で称徳天皇(女帝)をたぶらかして法王にまで出世したという弓削(ゆげの)道鏡からとったものだ。
錠と鍵
キチン質の骨片は、20世紀の初頭からオーストリアやフランスの昆虫分類学者によって、その存在は知られており、-部分類の指標に使われたことがある。日本でも第二次大戦後に、中根猛彦氏(元鹿児島大学教授)がこの骨片によってオオオサムシの分類を体系化し、骨片オサムシと通称されてきた。しかし、この骨片にどのような機能的な意味があるのかは誰も注目しなかった。
石川教授は以前からこのキチン骨片には、何か特別な機能があるのではないかと考えていた。そう考えると、ドウキョウオサムシの異様に大きな骨片は、交尾の邪魔になるだけにしか思えない。どうしてこんなものがついている必要があるのだろうか。
答えは意外と簡単だった。この新種のオサムシの雌を解剖したところ、骨片がすっぽりおさまるポケットが雌の交尾器にあることがわかった。しかも、顕微鏡の下で雌雄の交尾器を組象合わせてみると、間違いなく骨片とポケットは対応する。骨片は邪魔者ではなく、雌の交尾器のポケットを錠とすれば、まさに鍵のようにはまったのである。石川教授は、雌の膣にある、雄の交尾片に対応するポケットを「膣盲嚢(ちつもうのう)」と呼ぶことにした(図 3-1)。
すべてのオサムシが交尾片を持つわけではないが、日本のオオオサムシ亜属には、とくにさまざまな形の交尾片が存在する。交尾片の形によって、中根氏はオオオサムシ亜属をヒメオサムシ、オオオサムシ、ヤコンオサムシ、アオオサムシの4種に分けた。石川教授は、この四つの種を種群とし、クロオサムシ種群を加えてオオオサムシ亜属を5種群、15種に分類し直した。
それぞれの種群は、体の小さいタイプと大きなタイプとに分かれる。小さいタイプは、主に山地に分布している。色彩はおおむね、地味な茶色である。これを、「一次種」と呼ぶ。大きいタイプは主に平地に分布し、棲息域ははるかに広い。こちらを「二次種」と呼ぶ。
まだ日本列島が中国大陸とつながっていたころ、渡ってきたオオオサムシ亜属の祖先種が日本の各地に分布を広げた。この祖先型の状態をよく残しているのが一次種で、その後大型のものが現れ(二次種)、日本各地へと拡散していった(図4)と石川教授は考える。
このような方向性を持った形態の変化を、「形質傾斜」と呼ぶ。なぜ、進化にこのような方向性があるのかは、諸説はあっても定説はない。巨大な角で有名なオオツノジカは、性淘汰説で説明されたことで知られている。雌が大きな角の雄を好むので、しだいに雄の角が肥大化したというのである。剣歯虎は、餌のマストドンが大きくなるに従って牙が大きくなっていったのだ、と説明されている。剣歯虎は、牙をマストドンの頸椎にまで届かせて殺す。マストドンが大きくなれば、牙の大きい虎が有利になる。こうして、剣歯虎の牙も巨大化していったが、マストドンそのものが滅びたために、剣歯虎も絶滅したのだというのだ。
オサムシの交尾片の形状変化も、生存に有利なことが何かあったためなのだろうか。「生存の有利不利と、ペ二スの巨大化とが関係あるのかどうかはわからない。今の段階では、現実に交尾片やペニスの形が変わってきているとしかいいようがない」と石川教授は言う。
おそらく今から数千万~数百万年前、陸つづきだったアジア大陸から何種類かのオサムシが何回かにわたって日本列島にやってきた。この何千万年、何百万年、あるいは数万年の間に、日本列島はダイナミックな造山運動やさまざまな気候変動を体験してきた。ときには、あるところで渓谷が生まれ、氷河期に低地を覆っていたブナ・ミズナラ帯は、高温期には海抜2000mを超える稜線にまで達した。オサムシはごく少数の例外を除いて、後翅が退化していて飛べない。こうした環境の変動が、オサムシたちをそのつど地理的に隔離し、現在のような6属35種に及ぶ日本のオサムシ相を生んだのだ、と石川教授は考える。
大陸のオサムシに比べると日本のオサムシたちは、派手な分化はしていないが、35種のうちじつに29種は日本の固有種であり、カタツムリを食べるので有名なマイマイカブリ(写真7-3)のように、首も胴も長く、さらに長い顎を持った「世界でももっとも特異な形」のオサムシも生まれた。狭いながら日本は、立派な進化のショーケースであり、その進化は今でも現実に進んでいるにちがいない。
コラム〔交雑〕
オオオサムシ亜属のオサムシ同士は、基本的には同じ構造の交尾片を共通に持つ点で一つの進化の道すじにある。分布域を接していれば、交雑の起こる可能性も考えられるのではないだろうか。事実、そうした例がかなりあるのである。
たとえば、三重県北部の鈴鹿山地では東西にマヤサンオサムシとイワワキオサムシが境を接して分布している。マヤサンオサムシはアオオサムシ種群に属し、交尾片は長い釣針型をしている。イワワキオサムシの交尾片は、短く偏平な桃の実型である。当然ながら雌の膣盲嚢も、それぞれの雄に対応した形をしている。したがって、この両種類のオサムシは交尾器の形がほとんど合わない。にもかかわらず、この両種類が交尾している形跡が現に存在するのである。
雌のイワワキオサムシの膣盲嚢に、マヤサンオサムシ雄の交尾片が折れた状態で残っているケースが報告されている。むりやりはずそうとして、折れてしまったのだろう。なかには、複数の雄の交尾片を体内に残しているイワワキオサムシの雌もあった。交尾片を折られてしまった雄の運命がどうなったかは不明である。ひどいのになると、はずれないまま雄も雌も死んでしまったケースもある。
無理に交尾をしたとしても、そこに子供ができるかどうかはさだかではない。たとえできたとしても、一代かぎりの雑種であり、不妊種である可能性が高い。不妊でない子供ができるなら、生殖的な隔離が不完全で、まだ亜種のレベルにあると考えられる。
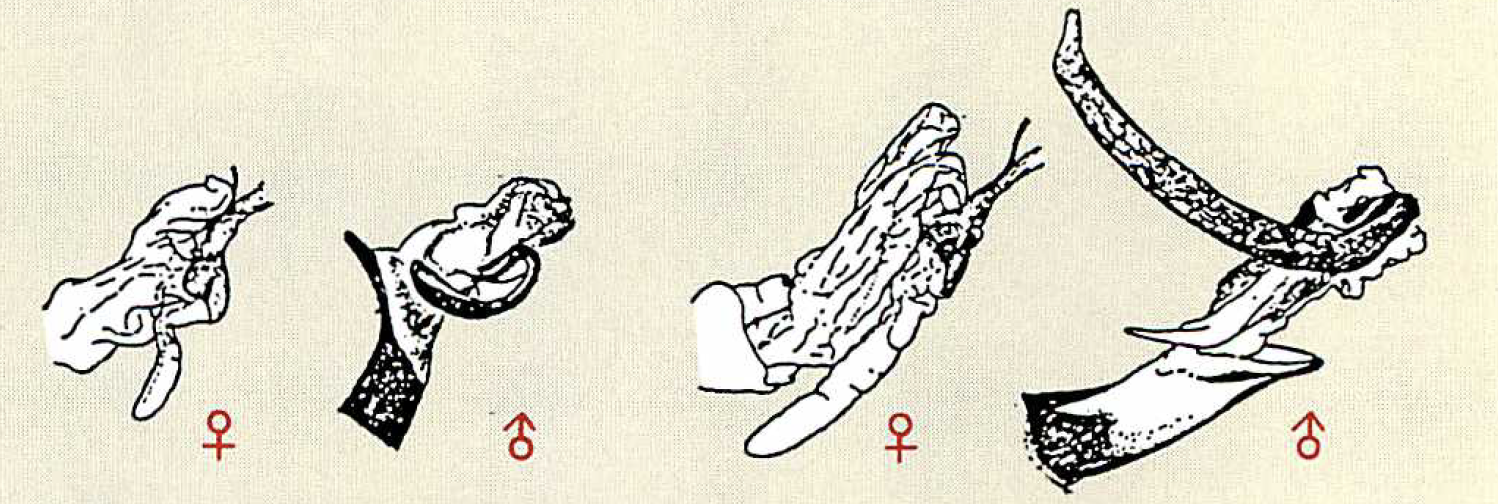
3-1 膣盲嚢と交尾片の関係図。アオオサムシ(左)と、ドウキョウオサムシ(右)。小指の形をした袋状のものが膣盲嚢
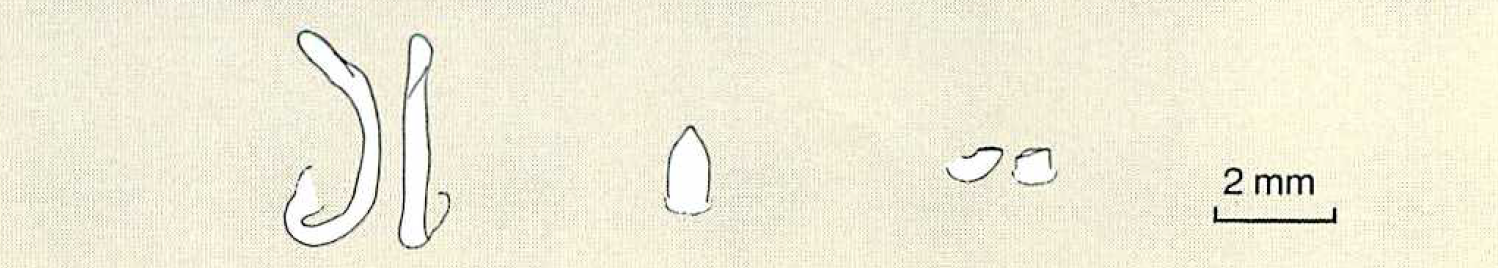
3-2 マヤサンオサムシ(左)とイワワキオサムシ(中)の交尾片。先端部が折れたイワワキオサムシの交尾片(右)久保田耕平(三重県立川越高校教諭)による
コラム〔骨片オサムシの進化と拡散〕
骨片で特徴づけられるオオオサムシ亜属の5種群のうち、アオオサムシ種群、ヤコンオサムシ種群、オオオサムシ種群の三つのタイプは、体の小さな祖形型からしだいに大きな種へと進化・拡散していったことが、交尾片と地理的分布との関係でよくわかる。アオオサムシ種群は静岡周辺、ヤコンオサムシ種群は紀伊半島、オオオサムシ種群は四国のそれぞれ山間部に一次種が分布している。
オーストリアのブロイニング(1894~1984)は、世界のオサムシを主に形態によって分類し、1600ページを超える大著『オサムシ・モノグラフ』を著した。日本のオサムシもおおむねブロイニングの分類によって、種や亜種が決められてきた。日本では一般的にこの3種群の一次種は、マヤサンオサムシの名で一つの種だと考えられていた。
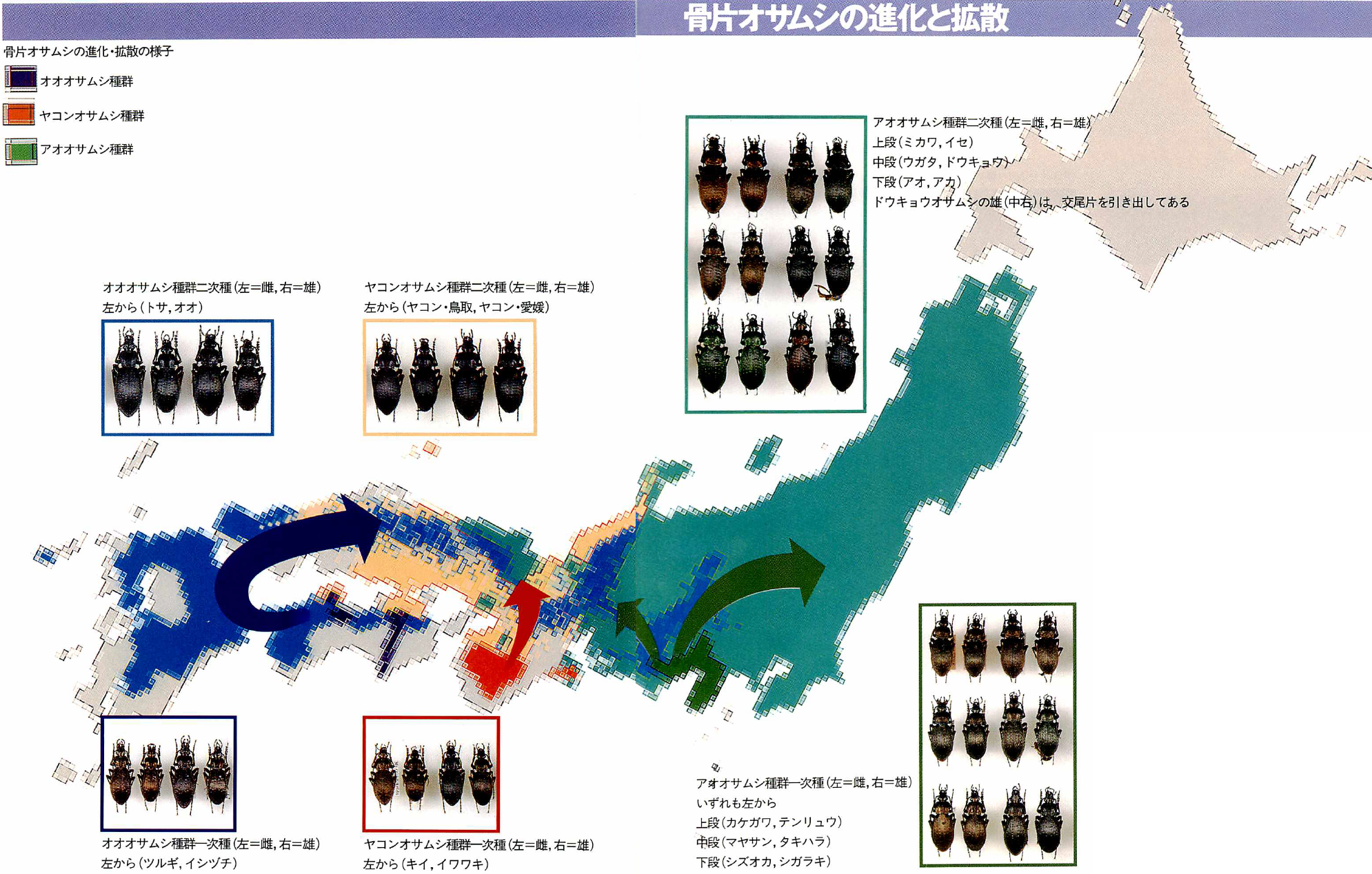
図4:骨片オサムシの進化と拡散
コラム〔系統樹で見る骨片オサムシの進化〕
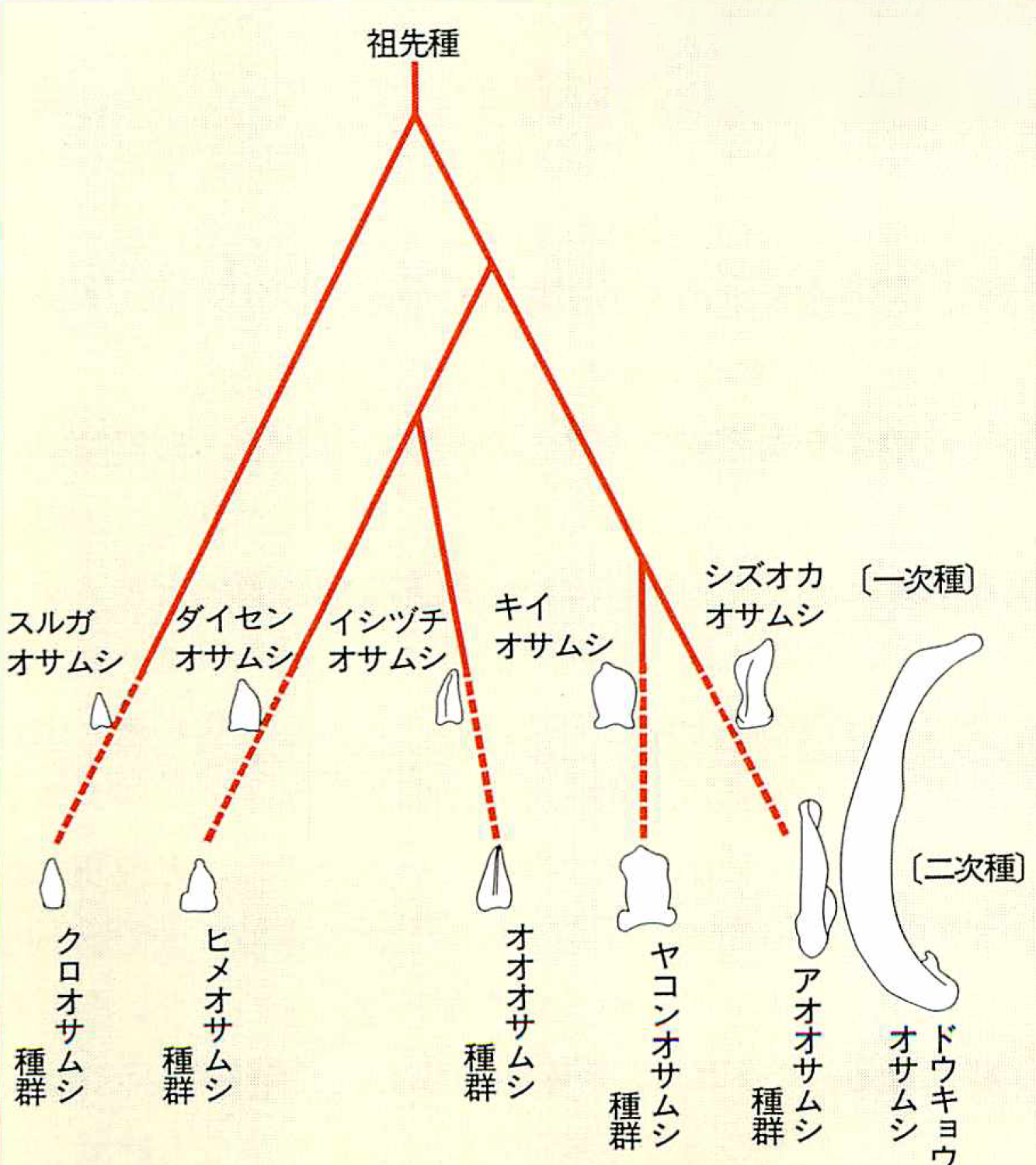
図5:系統樹とともに見るオオオサムシ亜属の交尾片の進化の様子
交尾片を持つオオオサムシ亜属の五つの種群の中で、三角形の交尾片を持つクロオサムシがその原型に近いと考えられる。三角形のタイプはヒメオサムシ、オオオサムシ、クロオサムシの3種群に分かれ、一方で桃の実型のヤコンオサムシ種群と、釣針型のアオオサムシ種群が別々に分岐していった、というのが、石川教授の考え方である。ドウキョウオサムシは、釣針型タイプの極致ということになる。
中部以北北海道まで分布しているクロオサムシ種群とヒメオサムシ種群は、交尾片の変異が少なく、体の大型化もあまり進んでいない。本州と北海道が分離する以前に、拡散を終えてしまった種群と考えられる。この2種群は地図上にはのせていない。
コラム〔交尾器による隔離機構〕
ある生物の分布が山や谷、川などの出現によって地理的に隔離されたとしよう。隔離された生物群は、それぞれの間で独自に突然変異が蓄積されていく。長い時間の間に、両者が相互に生殖不能な集団へと変化したとすれば、その生物は二つの新しい種へと分化したことになる。これが、地理的な隔離による種の進化である。
生殖的隔離には、いろいろなケースが考えられる。たとえばショウジョウバエは、雄が雌に対して行なう求愛行動が種によって特異的で、雌はこのパターンが自分の種のものでなければ、その雄との交尾に応じない。蝶では翅の色を雄が仲間の雌の識別に使っている。このように多くの種は、行動や色彩、あるいは雌の出す性フェロモンなどによって、交尾対象を識別しているのである。
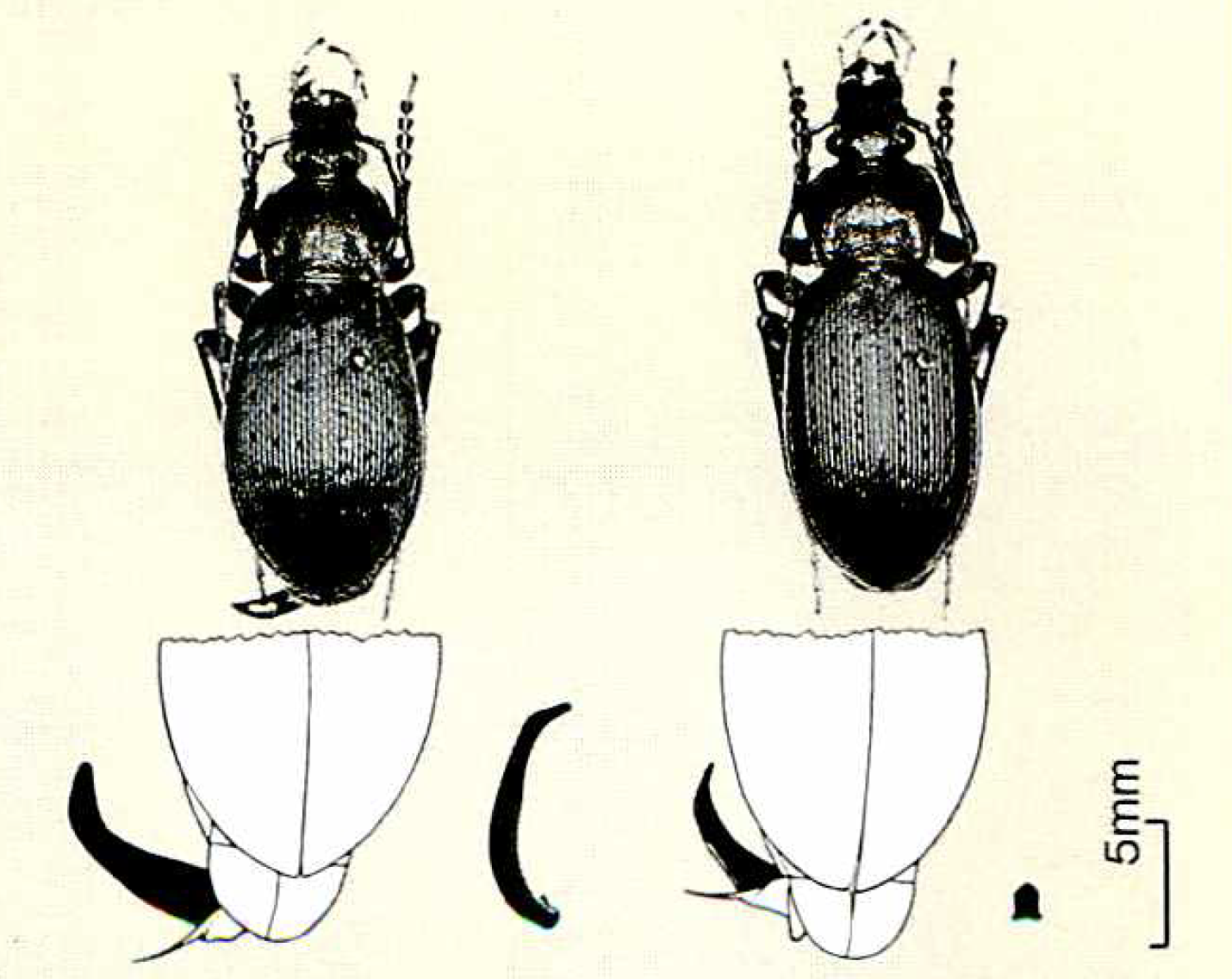
図6:ヤコンサオムシ(右)の雄とドウキョウオサムシ(左)の雄。外形はそっくりなのに、ペニスと交尾片はこんなに違う
行動でも色彩でも、それはある種の「信号」である。交尾のための信号が異なれば、たとえ外形が類似していてもその個体同士は交尾を行なわない。ところがオサムシの場合は、いささか異なっている。この虫は、同種の仲間を識別する信号をとくに交わさないらしい。石川教授によると、オサムシの雄は相手が異なる種のオサムシでも、あるいは同じ種の雄でも、大きささえ適当であれば相手かまわずマウントして交尾しようとする。仲間のオサムシの雄に乗って交尾できないことに気づき「おや失礼」と引き揚げる、といった光景がままあるのだという。
とすれば、このオサムシたちは、どのようにして交尾可能な対象を識別するのか。じつは識別するというのではなく、交尾器の形によって現実的に交尾が可能かどうかが決まってしまうらしい。形が合わなければ、結果的に雄と雌はまぐわえない。こうした形での生殖的隔離を「機械的隔離」と呼ぶが、これまで種の分化の要因としてあまり重要視されてこなかった。石川教授は、日本のオサムシにおいては、この機械的な隔離機構が、種の分化におおいにはたらいたと見る。
コラム〔東海地方におけるアオオサムシ種群の進化・拡散〕
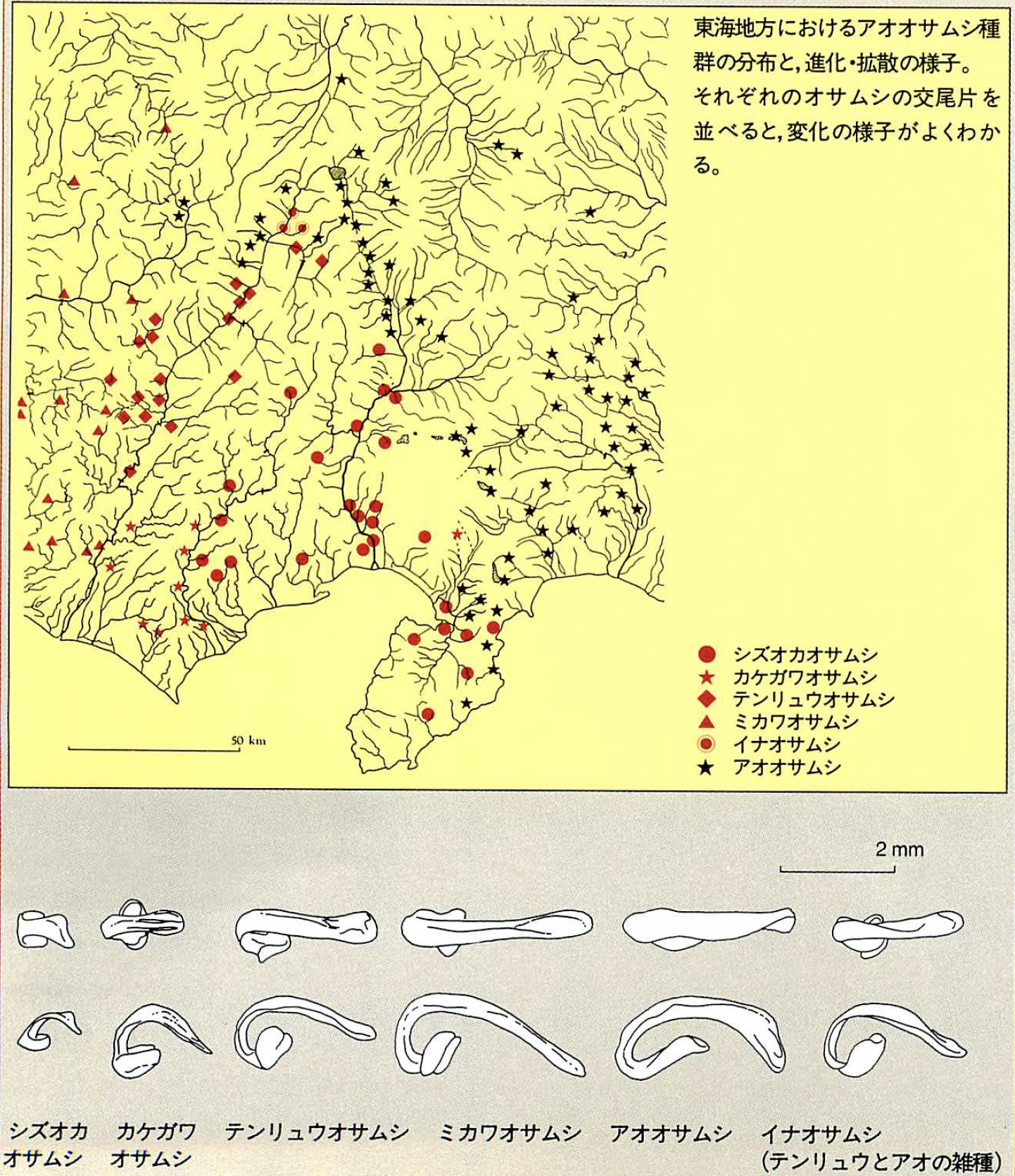
交尾片の形の変化は、剣歯虎の牙のように方向性を持っているように見える。この骨片の変異と棲息域との関係を見ていくと、オサムシの地理的な拡散の様子が浮かんでくる。たとえば、アオオサムシ種群に属しているシズオカオサムシ、カケガワオサムシ、テンリュウオサムシ、ミカワオサムシを例に見てみよう。このなかでは、シズオカオサムシがもっとも祖先的な形状を残しており、この種全体の祖先型にもっとも近いと考えられる。交尾片は小さなカギ型をしている。カケガワ、テンリュウとなるにつれて、このカギ型の先端が少しずつ伸びて全体的に大きくなる方向に変化していく。さらに、ミカワオサムシの先端がねじれた形のものがアオオサムシである。
シズオカオサムシは、伊豆半島を含めたほぼ静岡県全体に分布している。その西隣にカケガワオサムシ、そのまた西にミカワオサムシがおり、カケガワのすぐ北隣にはテンリュウオサムシがいる。交尾片の方向性からすれば、このオサムシの仲間は、シズオカ、カケガワ、テンリュウ、ミカワといった順に、図のように進化をしながら地理的に拡散していったと考えられるのである。
アオオサムシは、シズオカ、テンリュウ、ミカワとの境界から関東と東北の全域にわたる非常に広い棲息域を持っている。石川教授は「まだ、恐らくとしかいえないが」と断りながら、この種はカケガワオサムシとは別個にシズオカオサムシから分化し、急速に棲息域を拡大したと考えられるという。
コラム〔遺伝子でオサムシを見る〕
遺伝子DNAは、A(アデニン)、G(グアニン)、T(チミン)、C(シトシン)の四つの塩基がつらなった二重らせんの分子である。長い進化の時間の中で、遺伝子の文字列は突然変異によってある一定の割合で置き換わっている。置き換わりの割合は、ほぼその生き物の存在時間に比例するので、その割合の違いから生き物同士の遺伝的な距離を推定できる。つまり、遺伝子は分子の時計でもあるのだ。

図 7-1 クロカタビロオサムシ(長野・白馬山頂)。オサムシに近いこの仲間には飛べるものが多い。
この遺伝子時計は、いくつもの新しい事実を明らかにしてきている。たとえば、およそ3000万年前にさかのぼると考えられていたヒトとチンパンジー・ゴリラの分岐年代は、ずっと新しく500万年前になることが示された。シダはコケから進化してきたと考えられていたが、先にシダがあってコケは逆に退化したものであることを分子時計は示している。

図 7-2 ダイセツオサムシ(北海道・白雲岳)
オサムシが、どのようにしてさまざまな種に分かれていったのかも、遺伝子レベルで調べることが可能である。生命誌研究館では、オサムシのミトコンドリアDNAを使って、およそどのような年代に、それぞれのオサムシが分岐していったのかを調べる計画だ。

図 7-3 マイマイカプリ(山梨、韮山市内)。カタツムリを食べるので有名なこの昆虫も、オサムシの仲間。かじったカタツムリを消化液で混ぜている。
※所属などはすべて季刊「生命誌」掲載当時の情報です。














