

Special Story
バクテリアから生きものの基本を探る
進化というとよいものになっていくというプラスのイメージ、変異というと変わりものというマイナスのイメージで捉えられているようです。しかし、進化の原因は変異です。変異は、どのように起こり、どのようにして進化につながるのか。変異が起きても、うまく生きていけない個体ばかりで種が滅びてしまっては、元も子もありません。そこで「元本保証をしたうえで多様性を産み出す」という方法が、DNAそのものの中に入っているというのが、古澤理論の基本です。ちょっと面倒な概念も出てきますが、図を見ながら考えを追うと、考える面白さが味わえると思います。バクテリアだからこそできる実験結果も踏まえた論を皆さんも一緒に考えてください。
進化が起きるためには、まず変異がおきなければならないということは、ダーウィン以来言われてきたし、今では変異はDNAに起きる変化であることがわかっている。ところが、具体的な変異の発生様式から問い直した研究は行われていない。変異はランダムに入るのが大前提であり、誰もそれに疑問を抱かなかったからで、複製によってできた2つのDNA鎖には均等に変異が入ると信じられている。しかし、実際そうではない。変異は不均等に入るというのが私の結論であり、そこからは今までとは大きく違う進化の実態が見えてくる。大腸菌を例にしてDNAの複製を具体的に追いながら、変異の起き方のモデル2つを検討してみよう。
モデル1 連続鎖・不連続鎖モデル
DNAの二重らせんを作る2本の鎖は向きが反対であり、一方の鎖は連続的に複製されるのに対し、他方の鎖は部分的に複製した断片をつなぎ合わせる。不連続に複製されるほうは複雑な過程になるので、間違いの起こる率(変異率)が高くなるはずで、実際、連続鎖の10~100倍以上にもなる。つまり、複製の結果できた二重らせんの中にある変異の数は同じではなく、一方には多数の変異が入り、もう一方には変異が極めて少ないという不均衡が生じる。したがって、それらが入る2つの細胞も同じにはならないのだ。
じつはこれが進化にとって非常に大きなメリットとなる。変異の少ないほうで、元の親と同じ物(野生型)を確保しながら、その安全保障のうえで、変異率が高いほうでは、複製のたびに急速に増える変化をもとに「進化の実験」ができるからである。絶滅の危険を回避しながら、変異率をどんどん上げて、進化の速度を大きくできるのだ。私はこれを“元本保証の多様性創出”と呼んでいる。
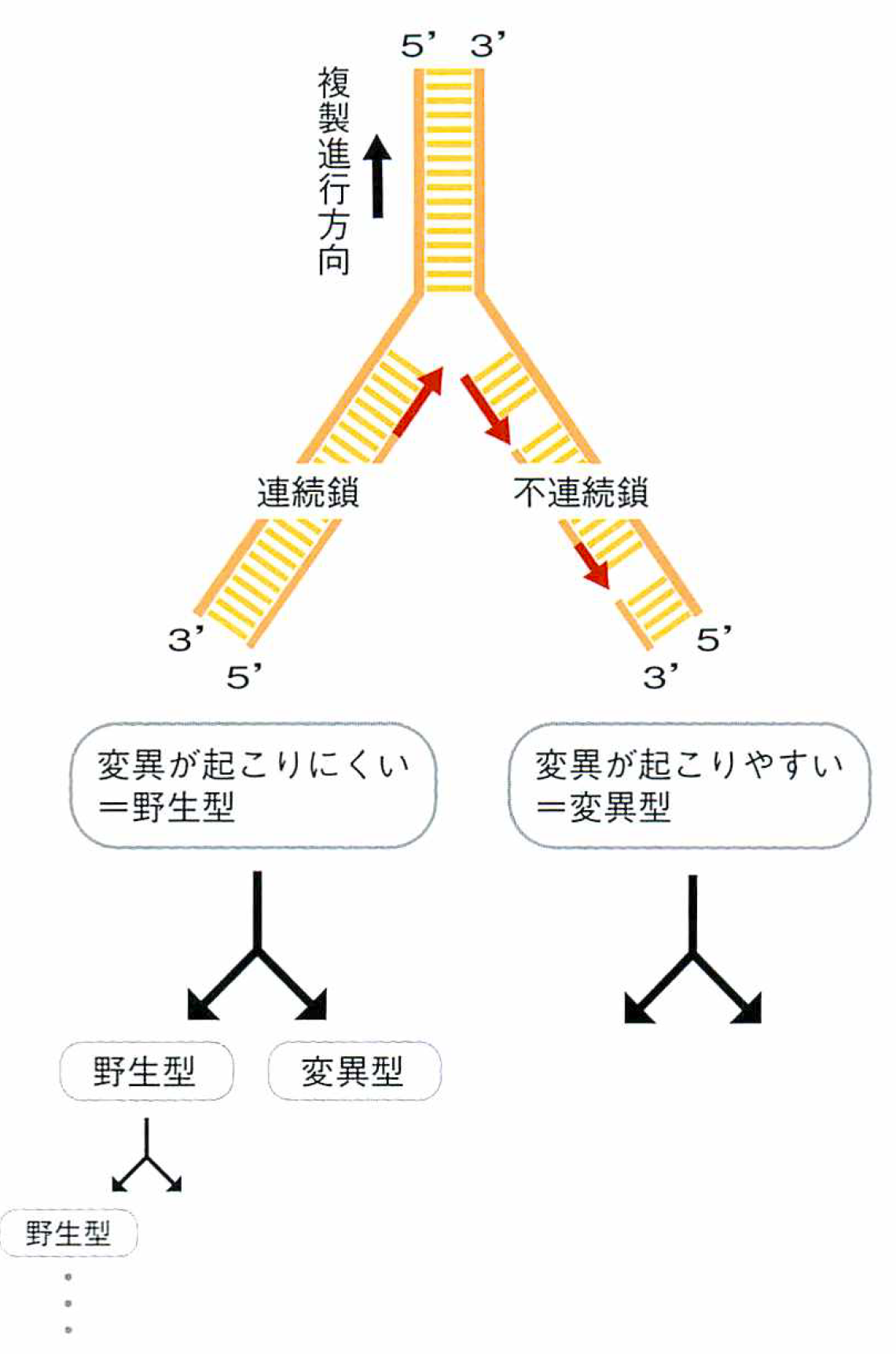
連続鎖・不連続鎖モデル
DNA鎖には、5’と3’と称される2つの末端があり、二重らせんを作る時には、それらがお互いに反対側に位置するように2本のDNA鎖が対を作る。複製は、この対をはずすのと同時に進められる。複製されるDNA鎖の伸長方向は、5’側から3’側と決まっているので、一方では対がはずれていくのに合わせて連続的に複製鎖の伸長が進められ(連続鎖)、もう一方では、対がある程度はずれるのを待って伸長開始を繰り返し、最終的に生じた断片をつなぎ合わせる方法で進められる(不連続鎖)。
連続鎖では変異の発生率が極めて低いため、野生型が保存されるのに対して、不連続鎖では変異率が高く、一度にたくさんの変異を導入した進化の実験が可能となる。
モデル2 混合複製酵素モデル
大腸菌DNAは環状であり、複製開始点から両方向に複製が始まり反対側で終わる。4個のDNA複製酵素が新しい鎖を作っていくわけだ。もしその4個の酵素のうちの1個が他より変異を起こしやすいと仮定すると、複製されたDNAのうち一方は変異がほとんどなく(野生型)、もう一方には億の変異が入ることになる。じつは大腸菌の複製酵素には、それが複製を間違えた時にその誤りを校正する酵素εが結合している。したがって、4個の複製酵素のうちのどれかにεが結合しない時には、その酵素が作ったDNAには変異がたくさん入っていることになるはずで、上に述べたようなことが起こり得る。そして、この場合も、変異が均等に入っていると考えた場合に比べて、はるかに多くの変異の入った鎖を作り出せるわけで、“元本保証の多様性創出”が可能となる。
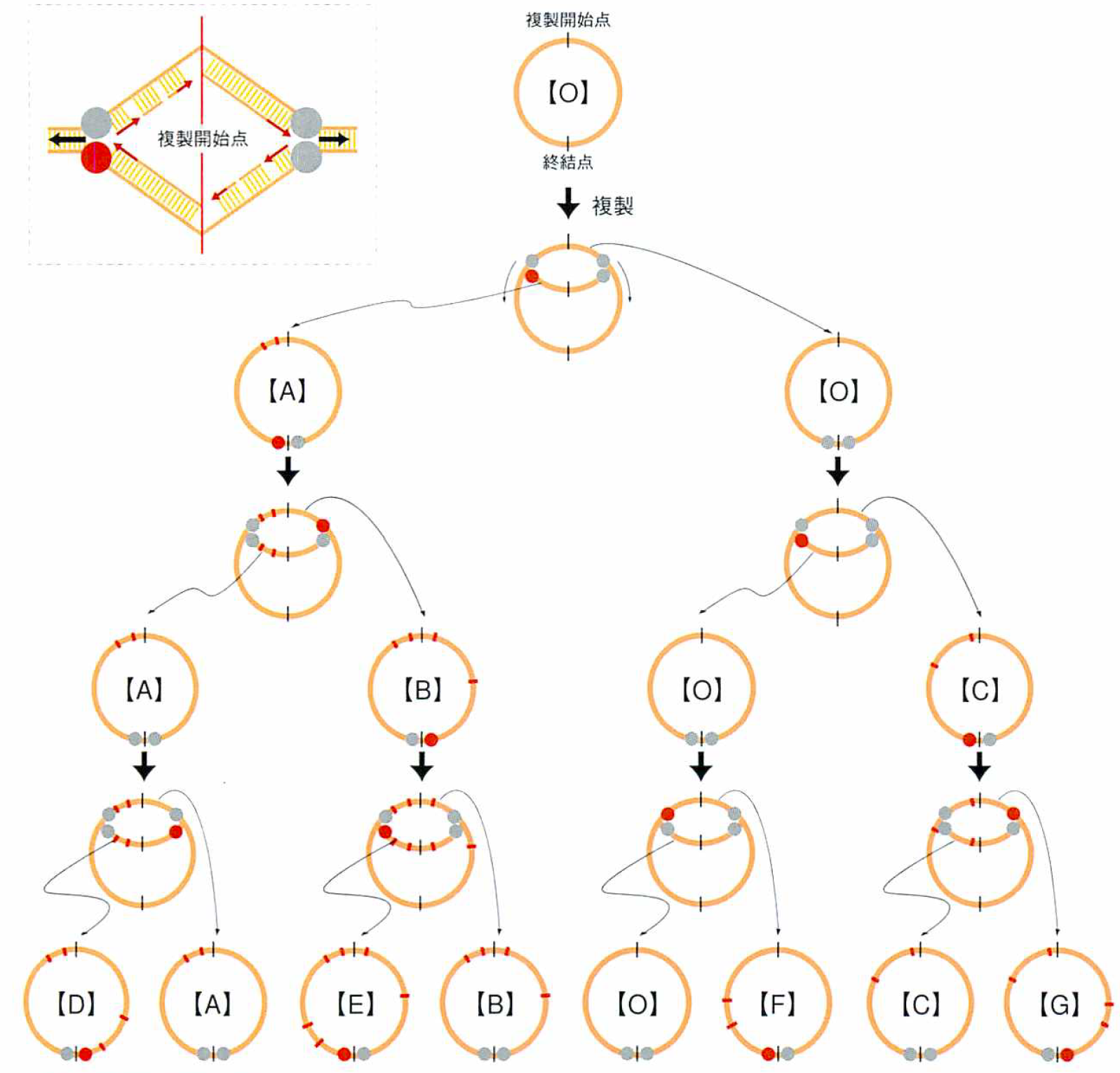
混合複製酵素モデル
大腸菌などの細菌のゲノムは環状をしている。複製開始の位置は決まっており(複製開始点)、そこから両側に複製が進められる。複製を進めるのは4個の複製酵素である(左上:ここでは連続、不連続といった複製の方法は考えない)。
通常、これらの酵素にはεという酵素が結合していて複製の精度を上げている。したがって、εとの結合がうまく果たせない複製酵素が混じっていると、複製鎖の一部に変異がたくさん入ることになる。このような酵素が一つだけであれば、複製鎖の一方は変異が多数入った変異型となり、もう一方は野生型のまま残るので、“元本保証の多様性創出”が可能となる(右下)。正常な複製酵素は(灰丸)、以上な複製酵素は(赤丸)、変異は(赤線)で表示した。
大腸菌での実験
じつは大腸菌に、校正の役割をする酵素εが通常の温度ではDNA複製酵素に結合しにくいという変異株がある。この株は、εがつかない複製酵素が適当量存在するような条件を作ることができるので、これを用いて実験してみたところ、4種類の抗生物質を過飽和に加えた培地でも平気で増える菌が短期間で得られた。つまり、変異率を大きく上げ、耐性菌への変化、いわば試験管の中での進化を、かつてない速さで起こすことができたといってよい。変異がランダムに入るのであれば、変異率を上げすぎると進化の前に全個体が死滅するはずだが、それが起きなかったのは、変異の発生が不均衡であることによって元本が保証されているからではないだろうか。
注目すべきは、この実験では、放射線や変異原物質などによって変異を起こしてはおらず、大腸菌が自然な形でDNAを複製し増殖する能力そのものの中にある進化の力を引き出したことだ。
不均衡で考える変異と進化
これまでは、「変異は集団に均等に分布する」と考えられてきた。そこで、変異率があまり高すぎると、すべての個体に多くの変異が入って全個体が死んでしまうことになるので、「変異の閾値がある(大まかにいってゲノムが1回複製するごとに1塩基が変わる程度)」とされてきた。したがって、進化には地質学的時間が必要だといわれてきたのだ。
しかし、変異が起きるDNAという分子はけっして対称構造をしているのではないし、その働き方も同じではない。「不均衡」。この事実に注目すると、変異の起き方、進化の実態が異なって見えてくるというのがこれまで述べてきたことだ。
ここまでは大腸菌で話しを進めてきたが、もっと複雑な生物でも不均衡効果は存在する。混合複製酵モデルの場合は完全な不均衡にはなりにくいが、連続鎖・不連続鎖モデルであれば、すべての個体で複製開始点が決まっており、性が存在すれば不均衡は生じる(詳細は省くが、複製の過程を追い、性による混合の際にはメンデルの法則を当てはめるとそれがわかる)。
カオスという概念を用いて生物の特徴を語ったカウフマンは、次のように説明する。変異率が低くて進化が起きない「定常状態」(熱力学の言葉を借りれば「凍結状態」)と変異率が高すぎて遺伝情報が攪乱し「融解」してしまう「カオスの海」との境目に「カオスの縁」があり、この微妙なバランスの場に生物があると。熱力学では、ものを平均化して扱うので、カウフマンはもちろん変位は均等に起きると考えている。しかし、不均衡変位を考えると、カオスの縁という特有の場を仮定せずとも、高頻度での変異が可能であり進化は起こりやすいものとして捉えられる。」あたかも沸騰するお湯の中に氷の小片が浮かんでいるような状態という表現をしてもよかろう。生物はカオスを乗り越えて存在し続けることのできるものなのだ。
環境が安定している現在、生物は休眠状態にあるように見えるが、生物は常に進化速度を調節して環境にしなやかに対応できる潜在能力を体内にもちつつ生きているのだと私は考えている。大腸菌という単細胞生物だからこそ、このような実験ができるわけで、ここに生物の面白さがある。
古澤 満(ふるさわ・みつる)
1932年生まれ。第一製薬株式会社特別参与。大阪市立大学理学部助教授、第一製薬分子生物研究室室長、取締役を経て現職。細胞生物学を専門とし、現在、進化の加速について研究を進めている。
.jpg)

.jpg)
.jpg)










