PERSPECTIVE
動物の腸内細菌
1. 消化を助ける細菌
私たちの毎日の生活には、食事が大切です。肉や魚、野菜、パンやご飯、いろいろな食べ物を食べることが健康的な食事だと考えられています。“You are what you eat”(あなたはあなたの食べた物でできている)という言葉がありますが、食べた物はどうやって、あなたをつくるのでしょうか。
食べ物を身体に取り込むためには、まず細胞が取り込めるように分解する必要があります。これを消化といい、食べ物を分解するのは消化酵素の役割です。ご飯やパンなどの「炭水化物」は唾液の消化酵素アミラーゼによって分解が始まります。一方、肉や魚などの「タンパク質」は胃液のペプシンで、「脂質」は胆汁と膵液のリパーゼで分解が始まり、最終的にそれぞれの最終単位となった「ブドウ糖」「アミノ酸」「脂肪酸」が小腸で吸収されます。主に野菜には、ビタミンや食物繊維などが含まれますが、ビタミンは小腸で吸収されるものの、食物繊維はほとんど分解されず大腸に送られます。
そこで、消化を語る上で忘れてはいけないのが「腸内細菌」の働きです。私たちは食物繊維を消化する酵素を持っていませんが、ヒトの腸にはおよそ1000種の腸内細菌が住みついており、私たちが自身の消化酵素だけでは分解できない食物繊維などを消化しているのです。また、腸内細菌はビタミンB群や消化管ホルモンなど多くの生理活性物質の合成にも関わっています。細菌たちのおかげで、日々の食事が私たちをつくることができるのです(図1)。
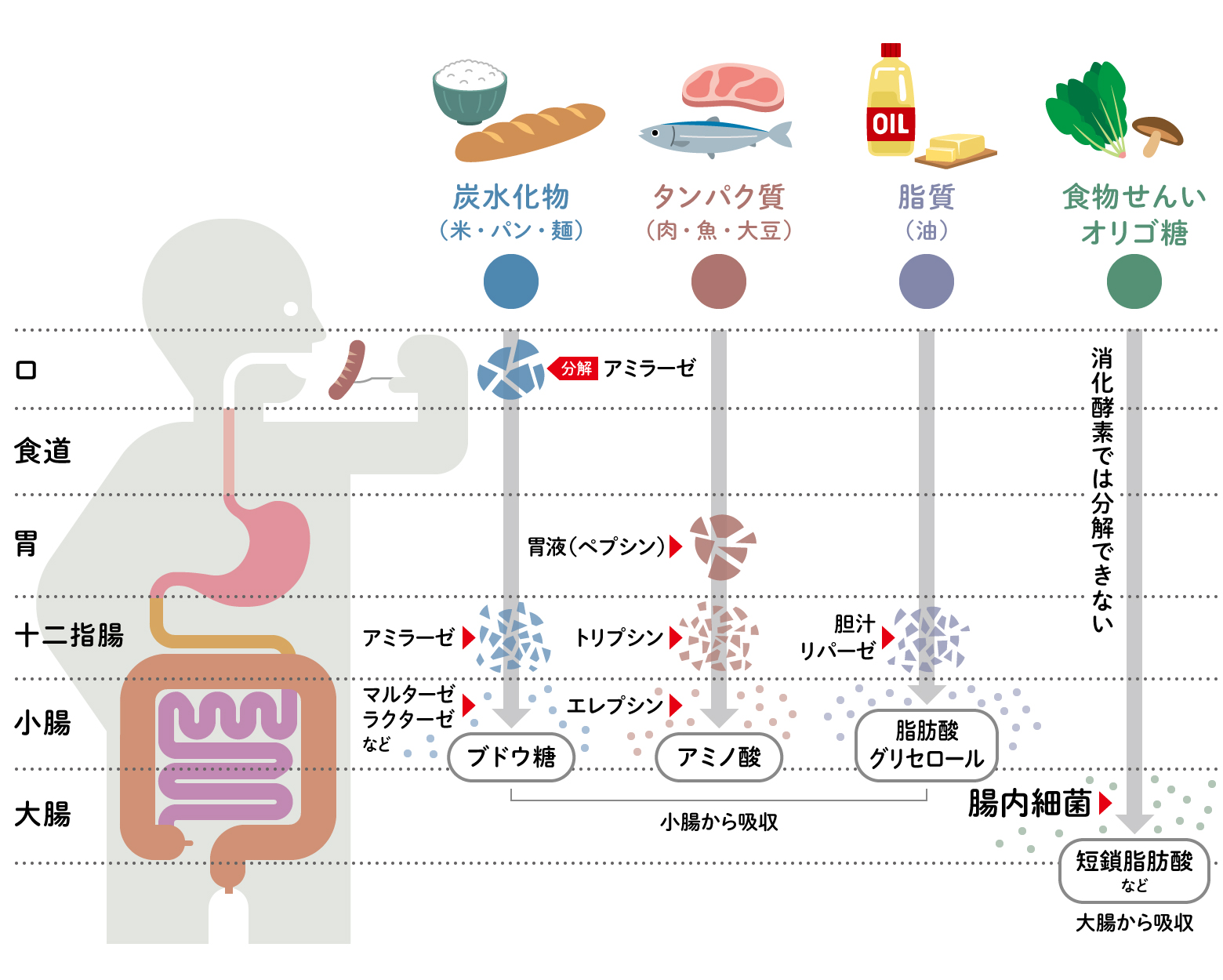
(図1) 食べ物を分解する消化酵素と腸内細菌
2. 腸内細菌とは
私たちの身体の皮膚、口の中、消化管の中には、たくさんの細菌が共生しています。そのうちの9割は小腸や大腸などの腸内にいる「腸内細菌」だと言われおり、全体でおよそ100兆個、1000以上の種類の細菌がいるとされます。口から入った細菌は多くの場合胃酸で殺菌されてしまいます。また肝臓から出てくる胆汁は界面活性作用を持ついわゆる洗剤なので、膜の弱い腸内細菌は溶かされてしまいます。ですからヒトも含めて生きものの腸内に住みつく腸内細菌は、一般に胃酸、胆汁酸耐性のあるものが定着しているわけです。胃を無事に通過した細菌は、小腸から大腸まで自分の住みやすい場所に集団で生息しており、そのうちの大多数は大腸に生息し、増殖を繰り返すことでその数を維持しています(図2)。たくさんの種の細菌が集団で生息している様子から「腸内細菌叢(そう)」と呼ばれ、この「腸内細菌叢」は年齢や食生活、動物種によって多様です。
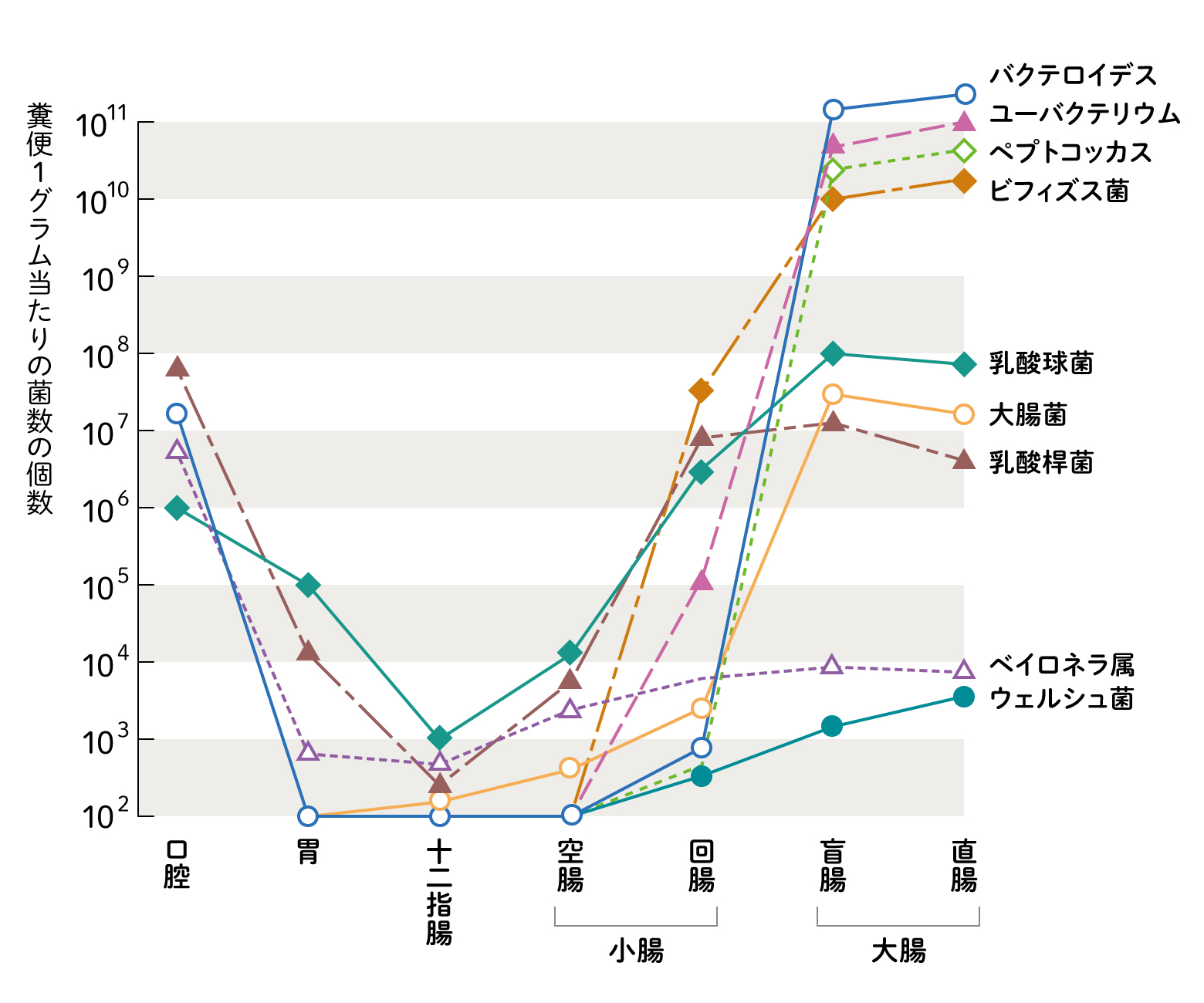
(図2) 消化管の各部位の菌叢
光岡知足, 腸内菌の世界, 叢文社, 1980年, p20 「図4 健康人の消化管各部位の菌叢」より一部改変
腸内細菌が、消化に深く関わっていることは前述の通りです。とくに植物を食べて暮らしている草食動物は、自身では食物繊維の成分であるセルロースやキシラン、ペクチンなどを分解する消化酵素を持っていないので、腸内細菌に頼って生きていることになります。ヒトのような雑食性の動物でも、デンプンの半分ちかくは難消化性であるため、じつは大腸まで到達してそこで腸内細菌の発酵作用を受けています。このようにして、草食や雑食の動物は、細菌が作り出す栄養素を腸管上皮細胞から取り込んで生活しているのです。
最近の研究からは、腸内細菌が腸管の粘膜組織を刺激して、自然免疫系の様々な分子の生産を促すことで、全身の免疫を活性化し、アレルギーや感染症への抵抗性などをもたらすことがわかってきました。さらに腸管の粘膜組織には、セロトニンやコレシストキニンなど神経伝達物質を創る細胞があり、一部は腸内細菌による刺激でこうしたホルモンの生産が促されることがわかっています。神経伝達物質は、血流を介して中枢に到達し、直接には食欲を調整したり消化管の運動を制御したりしますが、落ち着いた気持ちをもたらすような作用も示します。心の問題と腸の働き、そしてそれを促す腸内細菌という関係がいまでは「脳腸軸」という言葉で表現されるようになっています。このように腸内細菌は食物の消化を始め、免疫や心の健康までにも関係していることがわかってきており、私たちは生まれた時から一生の間、この腸内細菌と無関係で生活することはできません。
3. 食性と腸管
動物の中には、植物を食べるウシやウマなどの草食動物やライオンやネコなどの肉食の動物がいます。腸内細菌を身体の中に住まわせる場所としての腸管の構造は、生きものの食生活と大きく関わっています。特に草食動物は微生物の力を借りて消化を行っていますが、そのためには、消化器内で微生物の増殖を促すことと、発酵にかかる時間を稼ぐことが重要です。草食動物の腸管はこれらを前提として設計されています。消化管の長さが体長の10倍以上あることに加え、その一部が肥大して微生物の発酵タンクとしての役割を果たしているのです。偶蹄目のウシやキリンでは、胃の前にある「前胃」という3つの部屋に分かれた消化器官がその役割を果たし、前胃から何度も口へ戻し、咬み直しをしながら細かく分解する反芻(はんすう)によって消化を行います。一方、奇蹄目のウマやサイなどは、小腸の後ろにある長さ1メートルあまりの巨大な盲腸が発酵タンクの役割を担います。肉食性のイヌやライオンと比べると違いは明瞭です。肉食動物の消化管の長さは体長の3〜5倍程度と短く、食べたものはどんどん排出されます。消化を微生物に頼らず、自身の消化酵素で消化できるものしか食べないので、微生物をたくさん腸内に留める必要がないのです。一方、雑食性のヒトの消化管の長さは体長の7倍程度で、草食性と肉食性の中間の特徴を持っているといえます(図3)。
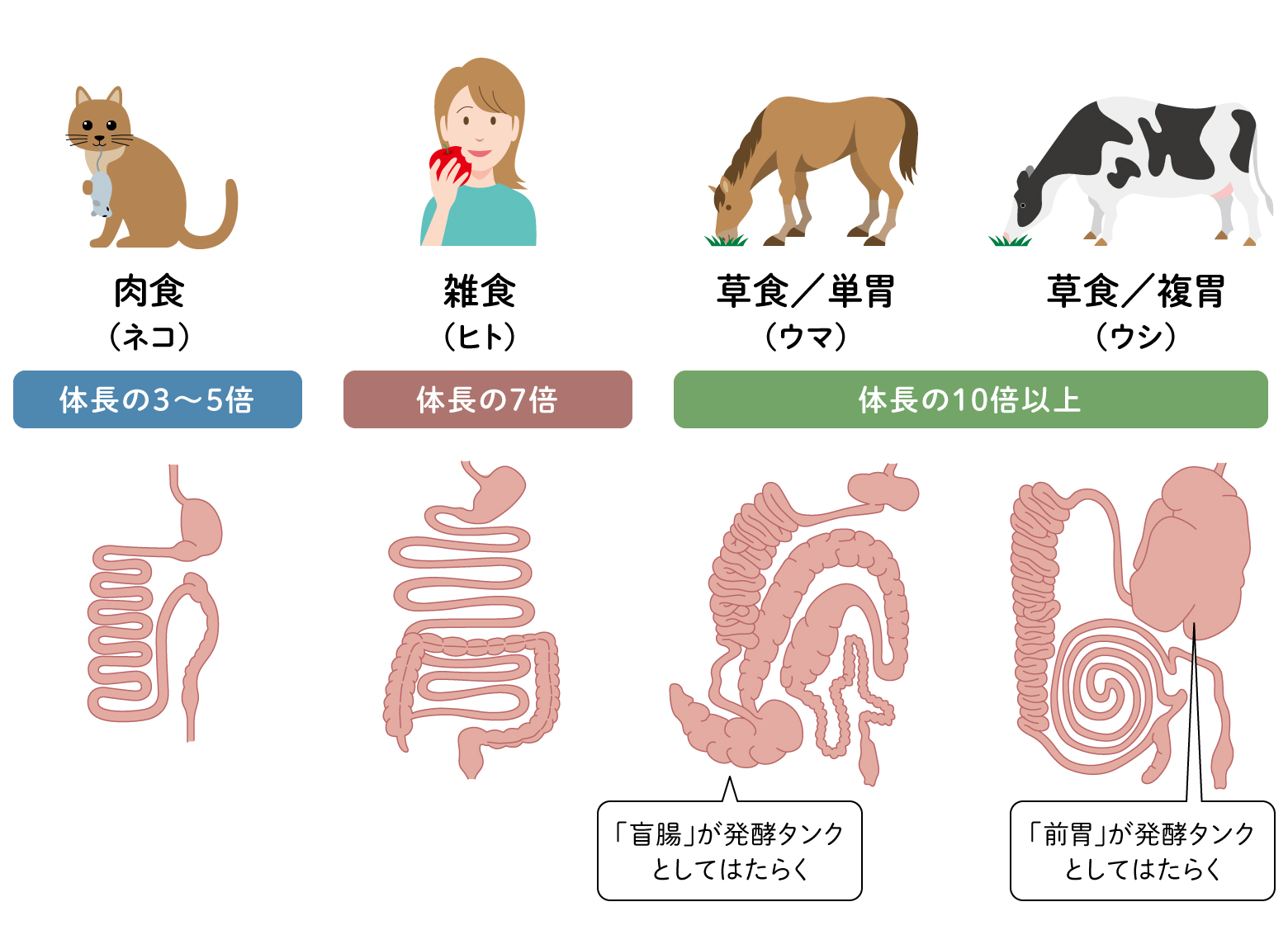
(図3)動物の腸管構造の比較
食生活の変化が腸管の構造に現れている例にはクマがあります。本来肉食動物のクマの系統は雑食にも適応しており、完全に肉食のホッキョクグマに対して、ツキノワグマなどのクロクマの仲間はドングリや果実、昆虫も食べる雑食性です。その腸管をみると小腸が長くなってきており、小腸を伸ばすことによって植物食に対応しようとしているようにもみえます。一方で竹・笹を食べるパンダもクマの仲間ですが、その消化管の長さはいまだに体長の4倍程度しかなく、その構造も植物食に対応した形状ではありません。しかし、クロクマの仲間と比べて大腸の長さが長くなりつつあるように見えるため、現在のところ大腸を伸ばして植物食に完全に適応できるように進化している途中かもしれません。このように哺乳類の消化管は食物と腸内細菌に合わせて進化しており、消化管に共生する微生物との関係性の歴史と、これから向かう方向が、その形状に表れているといえます(図4)。
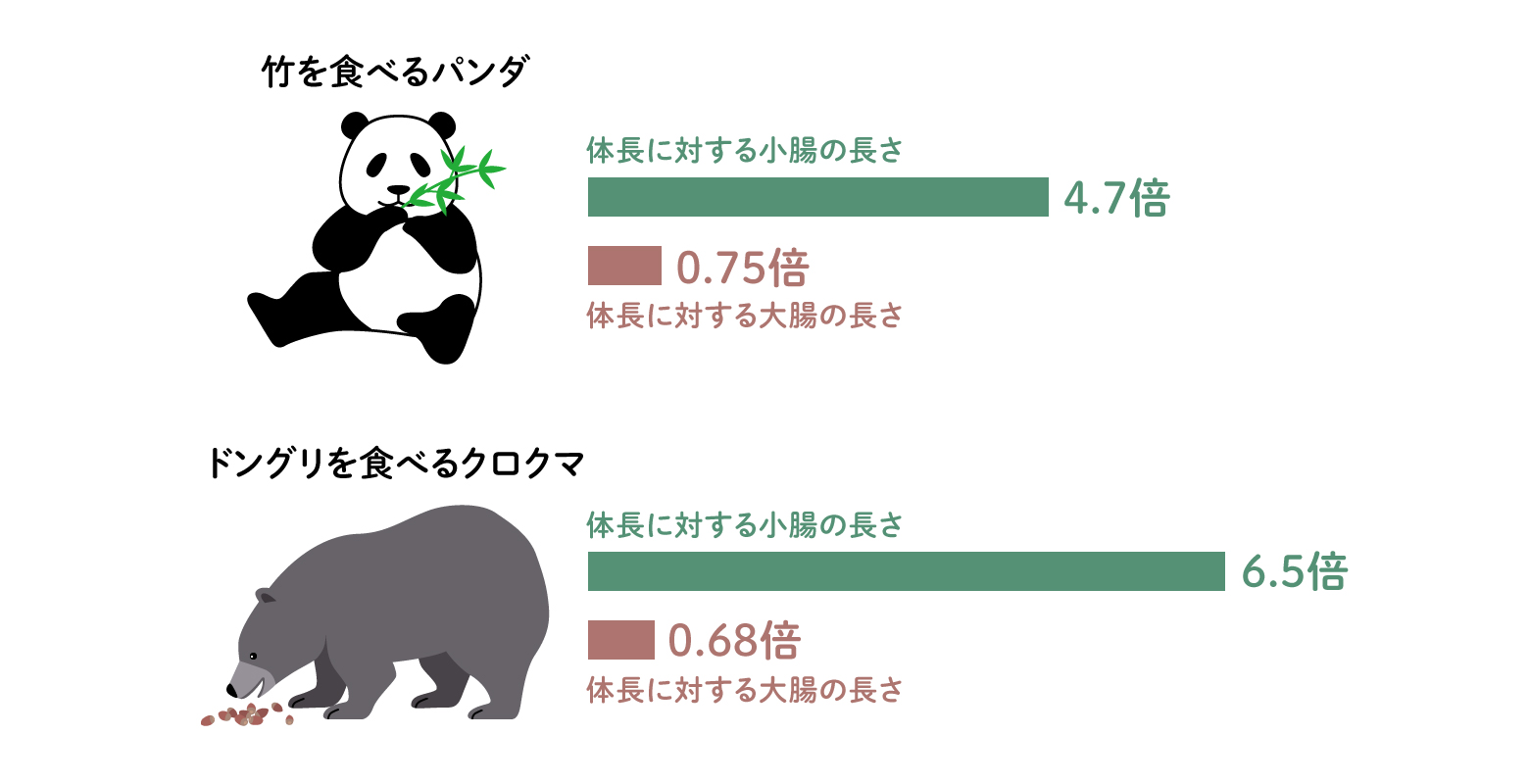
(図4) 食生活の変化と腸管の変化
4. 受け継がれる腸内細菌
腸内細菌叢は、その生きものの食性や年齢、腸管の場所によっても変化します。生活に必須の腸内細菌ですが、いったいどこから、どのように受け継がれるのでしょう。哺乳類の場合、体内にいる胎児は無菌状態で、分娩の時に初めて、母親の産道にいる乳酸菌、肛門のそばにいる大腸菌などに感染します。また卵から生まれる爬虫類や鳥類の場合、孵化したときに周辺の環境の細菌に感染します。哺乳類は、生まれたあと哺乳期間があり、母乳の栄養成分にはオリゴ糖が多く含まれていますが、実はヒトはその消化酵素を持っていません。腸内細菌が分解するのです。オリゴ糖を分解する細菌であるビフィズス菌は、乳児の細菌叢では主要な菌ですが、乳児は母親など世話をしてくれる人や入浴などを通して、ビフィズス菌を受け取ることがわかっています。そして、腸内細菌の種類が劇的に変わるのが離乳の時です。離乳食を食べ始めるようになると乳児特有の腸内細菌は消失して、成人と同じような腸内細菌叢が形成され始めます。
ヒトの場合は腸内細菌に消化を頼る程度はそれほど高くはないので、周囲の誰かから確率的水平的に腸内細菌が受け継がれるしくみでも問題ありません。一方で、生まれてすぐ大人と同じ食べ物を食べなくてはいけない動物では、その食べ物を分解する腸内細菌の受け渡しは非常に重要です。草食動物の場合、一番直接的な世代を超えた受け渡しは「食糞」です。たとえばコアラは母親が子育ての中で子どもにフン、それも腸内細菌の豊富な特殊なフンである「盲腸糞」を食べさせます。コアラの食べるユーカリには青酸化合物や高濃度のタンニンが入っており、多くの動物にとっては毒物となります。コアラは青酸やタンニンを分解する腸内細菌を母親から与えられて初めてユーカリを食べられる身体になるのです。同じような理由で、シマウマやキリン、霊長類などでも子どもによる食糞が知られています。鳥類でも、高山帯に住むライチョウは他の鳥のように孵化したあと親がエサを運んでくれません。そのため、孵化後から親と同じ高山植物を食べますが、高山植物には毒性を示す様々な二次代謝産物が含まれており、これらを分解するための腸内細菌を持つ必要があります。そこで、ライチョウの雛はふ化後3日から10日間ほど母鳥の排泄したフンを食べ、本能的に腸内細菌を取り込みます(図5)。
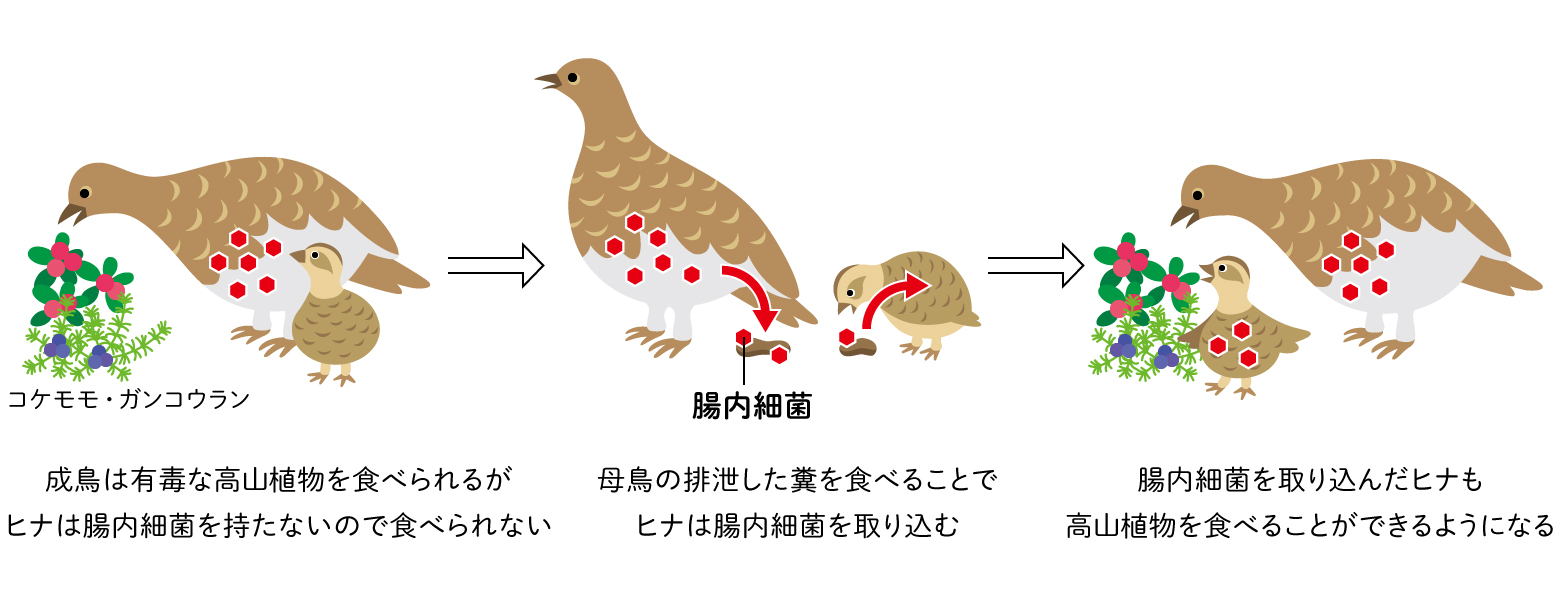
(図5) 食糞による腸内細菌の受け渡し
こうした動物の場合、たとえば怪我をしたときに、人間の治療と同じ感覚で抗生物質を投薬すると、消化に必須の腸内細菌が死んでしまうことがあるので、薬によって逆に生きていくことができなくなります。コアラやライチョウの雛の薬剤治療は困難が伴いますし、抗生物質を投与されたモルモットが、必死に仲間のフンを食べる姿をみていると、消化や解毒を腸内細菌に頼っている動物は、本能的に腸内細菌の重要性を知っていて、それを体内に取り込もうとする行動がプログラムされているようです。
5. 腸内細菌と共に歩んだヒト
私たちヒトに最も近いチンパンジーやゴリラは、森の植物の果実や葉などを主に食べ、もともと草食に適応しています。ですから彼らはたくさんの腸内細菌と共生するために長い大腸を持っています。一方でヒトの腸管はゴリラやチンパンジーなどと比べると大腸が小さくなっています。同じ祖先から進化する過程で、私たちヒトだけが今のような食生活スタイルに変化したからだと考えられます。
ヒトの祖先はアフリカの森で草食の生活を続けていましたが、800から1000万年前にゴリラと分岐し、それより遅い700から1000万年前にはチンパンジーから分岐したとされます。ヒトの直接の祖先だけが二足歩行になり300-400万年前には森を出て、その頃拡大していたサバンナに進出しました。その時に、従来の果物や葉が主体の森の食事から狩猟採集による生活に変わったことで、動物性の食物に依存する割合が大きくなったと考えられます。ゴリラはほぼ完全な草食者ですが、チンパンジーでは昆虫の摂食が多くなり、東アフリカのチンパンジーでは小動物を捕まえて食べる習性がよく観察されています。それでもチンパンジーの食物で動物性のものが占める割合は5%程度と見られています。現在の狩猟採集民の動物性食物の割合は50%から地域によっては90%に達します。
ゴリラやチンパンジーは立ち上がって歩くこともできますが、基本は四足歩行者です。彼らは長時間歩き続けることはなく、樹上にある食べ物を探しながら、食べて寝て森の中を移動します。ヒトの二足歩行は遅いですが、実は移動に関わるエネルギー消費はゴリラやチンパンジーの約半分ですむと推測されています。つまり同じ量の食物を食べても、そのエネルギーで移動できる距離は単純には2倍になるわけです。また体脂肪率に関しては、チンパンジーは5-7%程度しかありませんが、ヒトでは20%程度まであがりました。こうした代謝の変化により長距離を飲まず食わずで移動することができるようになったことが、サバンナでの狩猟採集生活に有利だったと考えられます。
それでは、腸内細菌の働きはどう変わっていったのでしょうか。ゴリラ、チンパンジー、ヒトに共通する腸内細菌はたくさんあります。一方、ヒトではビフィズス菌が圧倒的多数派ですが、ゴリラもチンパンジーもビフィズス菌は比較的少数派です。またゴリラやチンパンジーからは、ライチョウやコアラなどから見つかる乳酸桿菌の種類(リジラクトバシラス・アポデミ)や乳酸球菌(ストレプトコッカス・ガロリティカス)などがたくさん見つかりますが、ヒトで見つかることはありません。これらの乳酸菌は野生の植物に多く含まれているタンニンという消化を阻害する成分を分解する能力が高く、ヒトの食物からタンニンが次第に少なくなってきたためこうした細菌が必要でなくなった可能性があります。しかし、ビフィズス菌がヒトの腸内でなぜこれだけ多数なのか、その理由は実はまだわかっていません。私たちは、野生のニシローランドゴリラから、森の食生活に適応していたビフィズス菌を見つけましたが、ヒトの食生活がより肉食傾向の強い雑食へ変わる過程でヒトの中からは消えてしまったようです。
6. 腸内細菌のネットワーク
腸内細菌と共生するには宿主と腸内細菌の関係がうまく成り立っている必要があります。腸内細菌は、1000種に及ぶ集団ですから、1対1ではなく、1対1000という関係です。さらに1000種類の細菌はそれぞれの役割が異なり、その間でもやりとりがあります。例えば、もっぱらセルロースを分解して得られた糖から酢酸などの有機酸を作り出す細菌もいれば、難消化性デンプンを分解して有機酸をつくるものがいます。こうした有機酸を利用して別の有機酸に変えることで生きている者もいます。増殖に必要なアミノ酸はアンモニアから合成できますが、こうした細菌は、アンモニアをつくり出す細菌と手を組んでいます。 さまざまな細菌がお互いに連携して極めて複雑な代謝経路を腸のなかに作り上げているのです。それをクロスフィーディング(栄養共生)と呼びますが(図6)、このような栄養的な関係が1000種類の腸内細菌の間で何通りもあり、その組み合わせは膨大で、複雑なネットワークが広がっています。そのネットワークに登場する腸内細菌の種類やその組み合わせはヒトによって違います。しかし、細菌の種類や割合に違いはあるものの、実は違う細菌種でも等しくセルロースを分解できるというように機能で見た場合、ネットワーク全体の働きとしては大きな差はなくなります。つまり腸内細菌一個一個の種類が違っていても、そのこと自体に大きな意味はなく、全体として必要な働きが行われる腸内細菌のネットワークが構築されていることが一番大事なのです。
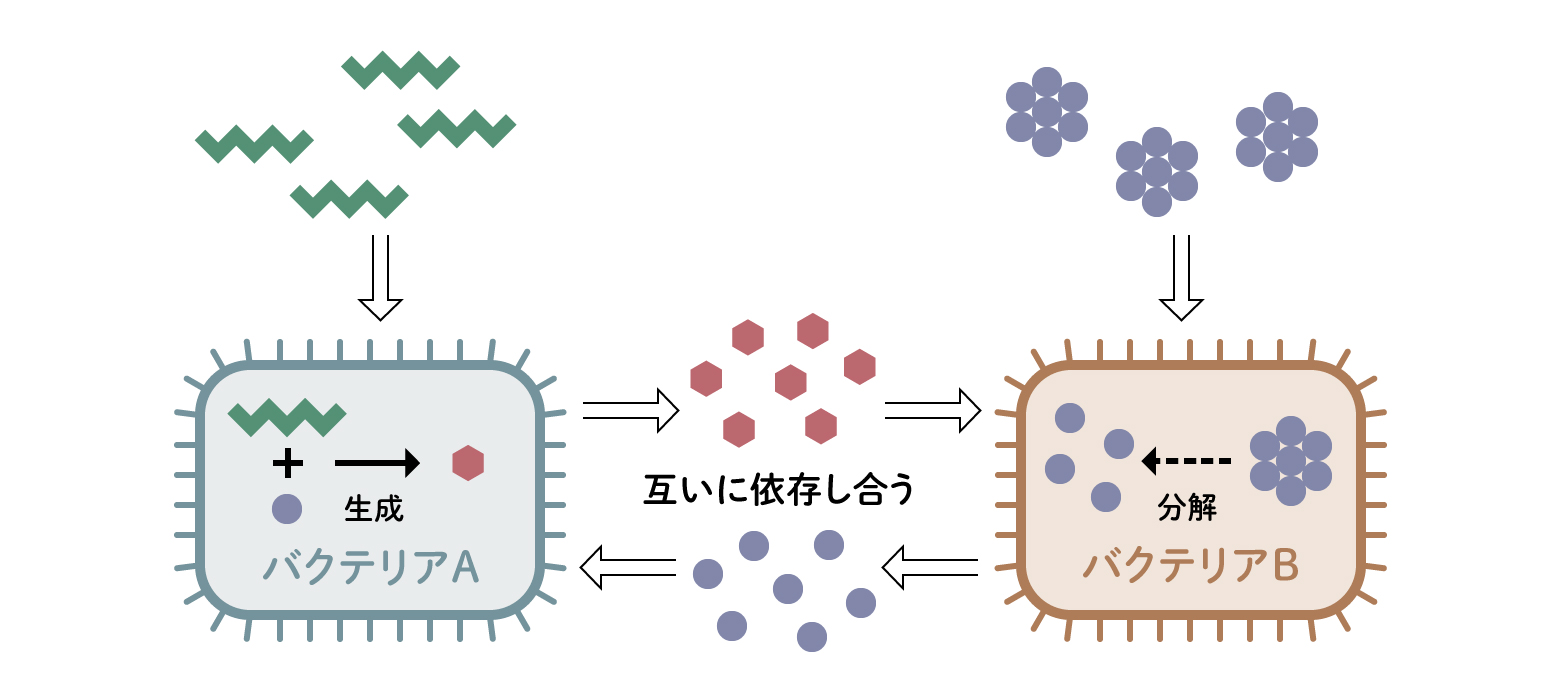
(図6) 腸内細菌のクロスフィーディング
7. フィールドと研究室を行ったり来たり
大学に就職してから30年あまり食品や製薬企業と腸内細菌や発酵食品の共同研究をしてきました。あるとき、なぜヒトでビフィズス菌がこれほど優勢なのか疑問を持つようになりました。ヒトに近いチンパンジー、そしてゴリラではどうなっているのか知りたくなったのです。動物園や研究所のチンパンジーやゴリラからはヒトと同じ種類のビフィズス菌を見つけることができましたが、あるとき市販ヨーグルトに使われている細菌をみつけました。あまり注意をしていなかったのですが、飼育されている彼らにはヒトの食べる野菜、さらにはヨーグルトも与えられていたのです。これでは、ゴリラ本来、チンパンジー本来のビフィズス菌を調べることにならないことに気付き、アフリカに渡って野生のチンパンジーの調査を始めました。これは腸内細菌学者として大きな挑戦でした。実は、動物園の動物ではなく本物の野生動物の腸内細菌の研究は誰もやっていなかったので、そもそも方法が未開発でした。実際、電気も蒸留水もないアフリカの熱帯雨林で腸内細菌を動物のフンから生きたまま分離し、維持することは現実的ではないと考えられていました。
動物のフンがあれば腸内細菌を調べることができますが、排泄されたフンが空気に触れないようにすることが重要です。腸内細菌の中には、嫌気性、つまり酸素に触れるとすぐ死んでしまうものがたくさん含まれており、こうした酸素に弱いものこそ、その動物にとって重要な役割を果たしていたり、他にはない独自のものだったりする可能性が高いのです。そのためアフリカの森の中では研究対象の動物にずっと張り付いて観察し続け、排泄を確認したら即座にフンを回収し、脱酸素剤と二酸化炭素を発生させる薬剤の入った容器に入れてキャンプ地に戻るまで保管しました。キャンプに着いてからフンを希釈し培地に塗抹してから同じように酸素のない容器内で培養しました。培養するには37℃に暖める必要がありますが、電気もない場所で一定の温度に容器を暖めるにはどうするか。最初は何も用意がなかったので人肌で暖めることから始め、お湯をつくって温度を維持することで対応しました。しかしこうした方法は小さな密閉式の試験管なら適用可能ですが、ペトリ皿を使う平板培地では不可能です。そこで3年目から手を温める携帯カイロと発泡スチロールの箱を日本から持ち込んで、培養箱を作り上げました。こうして一つひとつ手探りで方法をつくってきたのです(図7)。

(図7)ガボン共和国ムカラバドゥドゥ国立公園 森の中の「実験室」。ゴリラやゾウも訪れる。
アフリカのチンパンジーやゴリラから野生動物の腸内細菌研究を始めましたが、方法も開発し、あたらしい分野の扉を開いたと自負しています。ところで、同じ場所でアフリカゾウに出会うと、チンパンジーやゴリラと同じような食べ物を食べているゾウも調べてみたくなり、その次は近くをうろついているアカカワイノシシも、街をうろつくアフリカハゲコウも…という具合に、調査対象が広がっていきました(図8)。対象の動物種が広がる中で、培地の改良も進めなくてはならなくなりました。腸内細菌をスクリーニングするための培地の基本は、ヒトの感染症研究の中で確立された200年くらい前につくられたものですから、北里柴三郎やコッホが使っていた手法と基本的に同じもので、これらを改良しながら使っていることになります。もともとヒトやヒトのモデルである実験動物の腸内細菌研究のために開発された培地ですから、ヒト以外の動物に適用することには自ずと限界があります。
このようにして野生の霊長類から、偶蹄類、奇蹄類、さらには鳥類と、少しずつ対象を広げてきました。フィールド上に疑問の種が落ちていると、思わず拾ってしまう性格のようです。現在では、単なる研究だけではなく、ニホンライチョウ(Lagopus muta japonica)やヨウム(Psittacus erithacus)など絶滅危惧種の保全研究にも携わるようになりました。私は高校・大学・大学院を通じて山岳部に所属し、国内やヒマラヤで登攀活動をしてきましたから、もともと実験室の外の世界で研究活動をすることへの強い憧れがありました。いま、少しずつ確立してきた野生動物の腸内細菌研究を世界中のフィールドに適用できることに幸運を感じています。目の前の生きものから、未知の細菌をみつける研究をこれからも続けていきます。
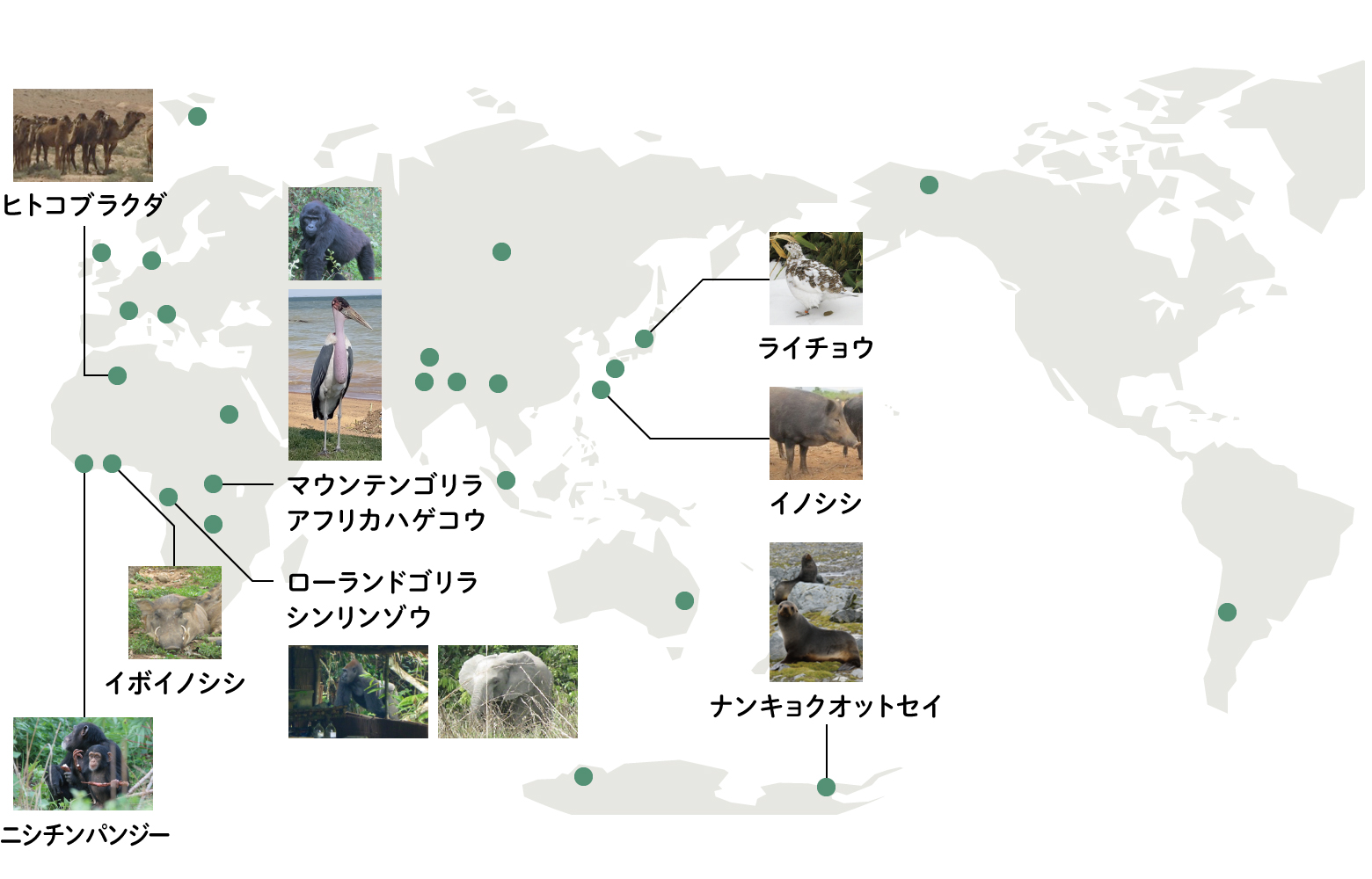
(図8)フィールドワークの主な調査地点と研究してきた野生動物たち

牛田一成
(うしだ・かずなり)
1954年12月兵庫県生まれ。専攻は、動物生理学/腸内細菌学。京都大学大学院農学研究科 博士後期課程修了(農学)。フランス政府国費留学生(フランス国立農学研究所 Theix 獣医畜産研究センター)をへて、京都府立大学農学部、同大学院生命環境科学研究科 教授/研究科長、中部大学応用生物学部教授/学部長 。(公財)日本モンキーセンター理事。(一社)京都大学学士山岳会理事。 環境省ライチョウ保護増殖検討会 委員。
「研究室サイト(外部サイト)」 「ヨウム保全プロジェクト(外部サイト)」














