RESEARCH
腸内細菌と宿主の肥満をつなぐ受容体
食物繊維は便通を良くするだけではない。腸内細菌が食物繊維を分解してできる短鎖脂肪酸が、エネルギーとしてだけでなく、私たち宿主側の細胞にある受容体にシグナルとして受け取られ、肥満を抑えていることがわかってきた。
1.腸内細菌と宿主の代謝疾患
腸内細菌の重要性は以前から指摘されていたが、その大半が嫌気性の細菌であり培養が難しいため、詳しい機能については明らかにされていなかった。ところが近年、ゲノム科学技術が進歩し、メタゲノム解析、マイクロバイオーム解析、16S rRNA系統解析などによって、培養せずにゲノムや代謝の解析が出来るようになり、腸内細菌の研究は大きく進んだ。その結果、腸内細菌と宿主のエネルギー調節や栄養摂取・免疫機能などとの関わり、細菌叢の構成変化と肥満や糖尿病・免疫疾患などとの直接的な結びつきが、科学的に証明されるにようになった。
中でも興味深い成果として、2006年のジェフリー・ゴードン博士らが明らかにした腸内細菌叢と肥満との関わりがある。肥満マウスと正常マウスの腸内細菌叢の組成を16S rRNA系統解析によって比べたところ、特定の細菌種に違いがみられた。そこで、肥満マウスの腸内細菌叢を正常マウスに移植したところ、体脂肪量が増えたのである。ヒトでも、肥満者と正常体重者との間で腸内細菌叢の構成比に違いがあることがわかった。ヨーロッパと中国における2型糖尿病に関する研究からは、人種や食事の違いに関わらず、全ての2型糖尿病患者に特定の腸内細菌種の変化がみられた。
2.腸内細菌の主要代謝産物 −短鎖脂肪酸−
これらの成果を受けて、研究の焦点は、腸内細菌の何がどのように宿主に作用しているのか、その分子実体である宿主側の受容体は何かということに移った。そこでは、腸内細菌自身だけでなく、それが作り出す代謝産物が鍵になっているのではないかという予測が立てられた。
腸内細菌の主な代謝産物は、食物繊維などを分解してできた酢酸、酪酸、プロピオン酸などの短鎖脂肪酸である(図1)。これらは宿主のエネルギー源としてだけでなく、細胞膜上にある短鎖脂肪酸受容体の活性化やヒストン脱アセチル化酵素の阻害によるエピゲノミックな作用をすることが見出された。
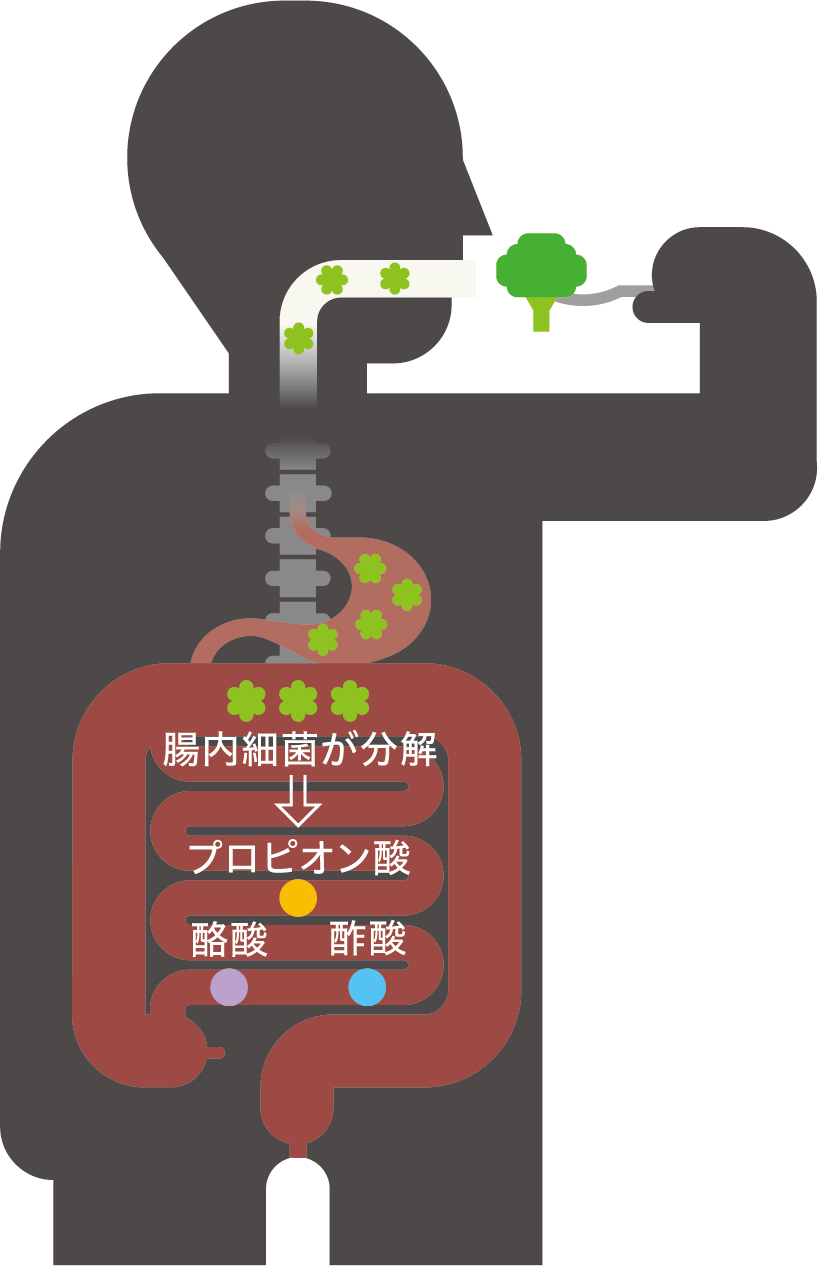
(図1) 腸内細菌が食物繊維を分解し、短鎖脂肪酸を産生する。
3.短鎖脂肪酸の受容体 –GPR41と43−
私たちはこれまで、各種の脂肪酸受容体の機能を解析してきた。それらは、7回膜貫通型受容体であるG蛋白共役型受容体(GPCR)に属し、遊離脂肪酸を受け取る細胞膜上にある受容体である。その中には、短鎖脂肪酸受容体GPR41とGPR43もある。そこで、これら2つの受容体が腸内細菌と宿主の機能をつなぐ分子となるのではないかと考えて研究を始めた。
GPR41は主にプロピオン酸と酪酸によって、GPR43は主に酢酸とプロピオン酸によって活性化され、それは循環血液中の短鎖脂肪酸濃度でも十分に起きることがわかっている。私たちは独自の発現解析によって、これら受容体が末梢の特定臓器で多く発現していることを見出している。そこで、腸内細菌が産生する短鎖脂肪酸は、腸内だけではなく、血流を介して末梢組織にある短鎖脂肪酸受容体に作用し、宿主の様々な生理機能に影響するのではないかという予測を立て、実験を行った。
4.エネルギー消費を促すGPR41
組織での発現を詳細に解析した結果、GPR41は特に交感神経節に多く発現していることがわかった(図2)。

(図2) GPR41の発現場所
GPR41のmRNAの局在(赤部)を調べたところ、交感神経節(矢尻)に多く見られた。
I.Kimura, et al., PNAS, May10, 8030-8035 (2011) より改変
そこでGpr41遺伝子が欠損したマウスを作出し、交感神経節でのGPR41の生理機能を詳細に調べた。すると、このマウスの心拍数は低下し(図3)、神経伝達物質であるノルアドレナリンの分泌量は通常よりも減っていた(図4)。
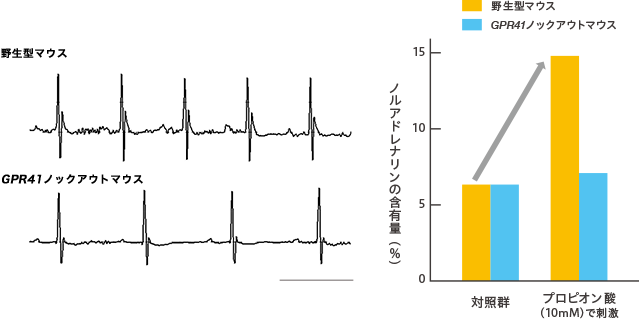
(図3)(左) 心拍数の変化
GPR41ノックアウトマウスでは、野生型マウスと比べて、心拍数が低下した。スケールバー:0.1秒。
(図4)(右) ノルアドレナリン分泌量の変化
プロピオン酸で刺激したところ、交感神経細胞からのノルアドレナリンの分泌量が増えた。一方、プロピオン酸で刺激しても、GPR41ノックアウトマウスでは増えなかった。
D.Inoue, et al., FEBS Lett., 586, 1547-1554 (2012)より改変
また、交感神経細胞を用いた実験において、短鎖脂肪酸によって刺激したところ、GPR41を介してノルアドレナリンの分泌が促された。これら短鎖脂肪酸によるGPR41を介した交感神経の活性化は、体温や酸素消費量を指標とした体全体のエネルギー消費量の上昇にも反映された(図5)。
ここから、GPR41は短鎖脂肪酸を介して交感神経系のコントロールにより、エネルギーバランスを一定に保つセンサーとしての役割を果たしていることがわかった。私たち自身が消化して得られるブドウ糖や脂肪酸などに加え、腸内細菌が産生する短鎖脂肪酸も宿主のエネルギー源であることが知られている。今回、短鎖脂肪酸はそれに加えて、宿主のエネルギー恒常性を保つ役割りをしていることがわかった。つまり、食べ過ぎても短鎖脂肪酸の血中濃度が上昇してGPR41を活性化し、交感神経を刺激することでエネルギー消費を高め、肥満が抑えられるのである。
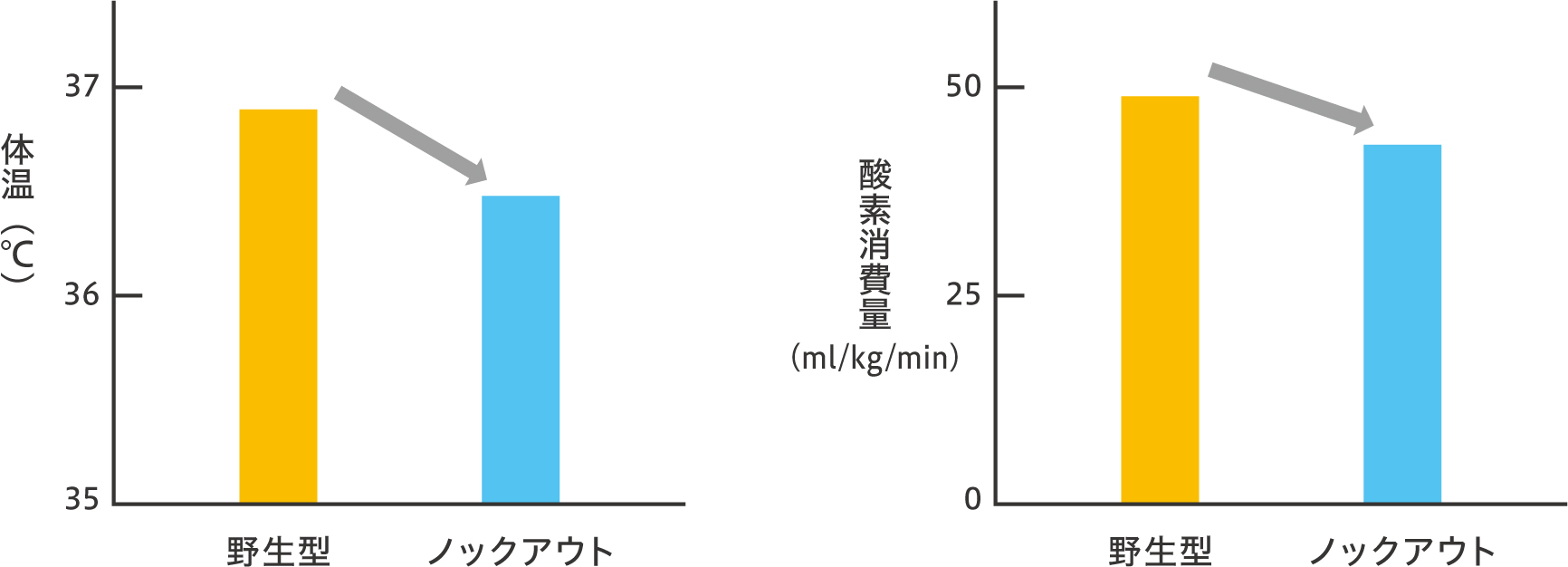
(図5) 体温と酸素消費量の変化
GPR41ノックアウトマウスでは、野生型マウスと比べて、体温(左図)と酸素消費量(右図)が低下した。
I.Kimura, et al., PNAS, May10, 8030-8035 (2011) より改変
5.脂肪蓄積を抑えるGPR43
一方、GPR43は特に白色脂肪組織に多く発現していた(図6)。そこでGpr43遺伝子が欠損したマウスを作出したところ、体重と脂肪重量が増し肥満傾向を示した(図7)。
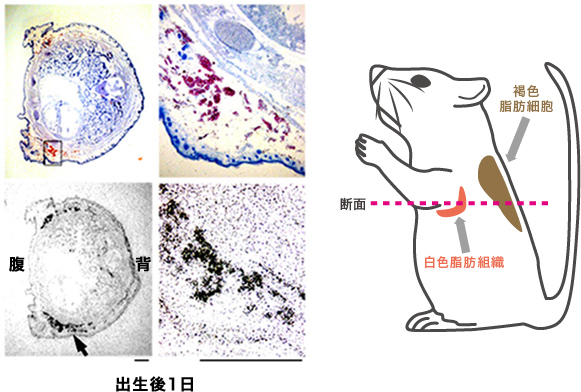
(図6) GPR43の発現場所
GPR43のmRNAの局在(上図の赤部)を調べたところ、白色脂肪組織(下図の黒部)に多く見られた。スケールバー:1mm。
I.Kimura, et al., Nat. Comm., May 7, 1-12 (2013) より改変
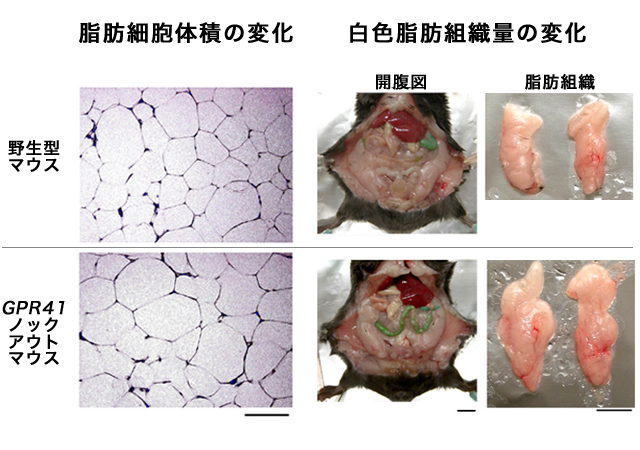
(図7) 白色脂肪組織量と脂肪細胞体積の変化
高脂肪食を与えた場合、GPR43ノックアウトマウスでは、野生型マウスと比べて、脂肪細胞の体積も増加した(左図)。スケールバー:0.1mm。 それにともなって、白色脂肪組織の量が増加した(右図)。スケールバー:1cm。
I.Kimura, et al., Nat. Comm., May 7, 1-12 (2013) より改変
逆に、白色脂肪組織だけにGPR43を過剰に発現させたGpr43トランスジェニックマウスでは痩せの傾向が見られた(図8)。無菌状態にしたり、抗生物質処置による腸内細菌の死滅を試みたところ、これらの代謝機能の変化は見られなかったので、GPR43を活性化する短鎖脂肪酸の供給源は、腸内細菌であると考えてよいだろう。
さらに詳しい分子メカニズムを調べたところ、GPR43からのシグナルは、脂肪細胞のインスリンシグナルだけを抑えることがわかった(図9)。
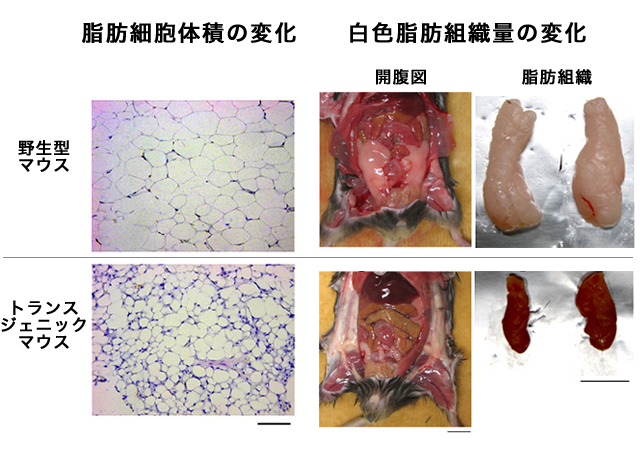
(図8) 白色脂肪組織量と脂肪細胞体積の変化
高脂肪食を与えた場合、GPR43を高発現させたトランスジェニックマウスでは、野生型マウスと比べて、脂肪細胞の体積も減少した(左図)。スケールバー:0.1mm。それにともなって、白色脂肪組織の量が減少した(右図)。スケールバー:1cm。
I.Kimura, et al., Nat. Comm., May 7, 1-12 (2013) より改変
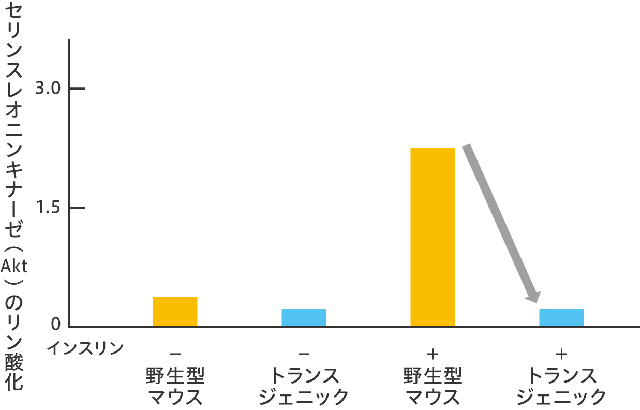
(図9) 白色脂肪組織でのインスリン作用の変化
GPR43を高発現させたトランスジェニックマウスでは、インスリンで刺激した場合も、野生型マウスと比べて、その作用が抑制された。その結果、白色脂肪組織においてのみ、セリンスレオニンキナーゼのリン酸化が抑制された。
I.Kimura, et al., Nat. Comm., May 7, 1-12 (2013) より改変
つまり、食べ過ぎてブドウ糖や脂肪酸などのエネルギーが過剰にあっても、GPR43が活性化されることによって、エネルギーの白色脂肪組織への取り込みを抑制し、脂肪の蓄積を抑えているのである。こうして、脂肪細胞の過剰な増加とそれによる全身のインスリン感受性の低下を伴う悪性肥満を抑えているのである。
6.さらに絡み合う、腸内細菌と宿主
私たちは、腸内細菌が産生する短鎖脂肪酸を、宿主が短鎖脂肪酸受容体GPR41と43を介して感知し、エネルギー恒常性を維持していることを明らかにした(図10)。特に、GPR41は交感神経系を、GPR43は白色脂肪組織におけるインスリンシグナルをコントロールしていることが明らかになった。
実は、GPR41と43は脾臓やリンパ節など免疫系組織にも発現しており、それらのエネルギー調節との関わりなど新たな課題が生まれてきた。また、他にも短鎖脂肪酸受容体が見つかってきており、それら受容体全体の相互作用を調べることが、腸内細菌と宿主の代謝機構の関わりを解く鍵になるだろうと考えている。
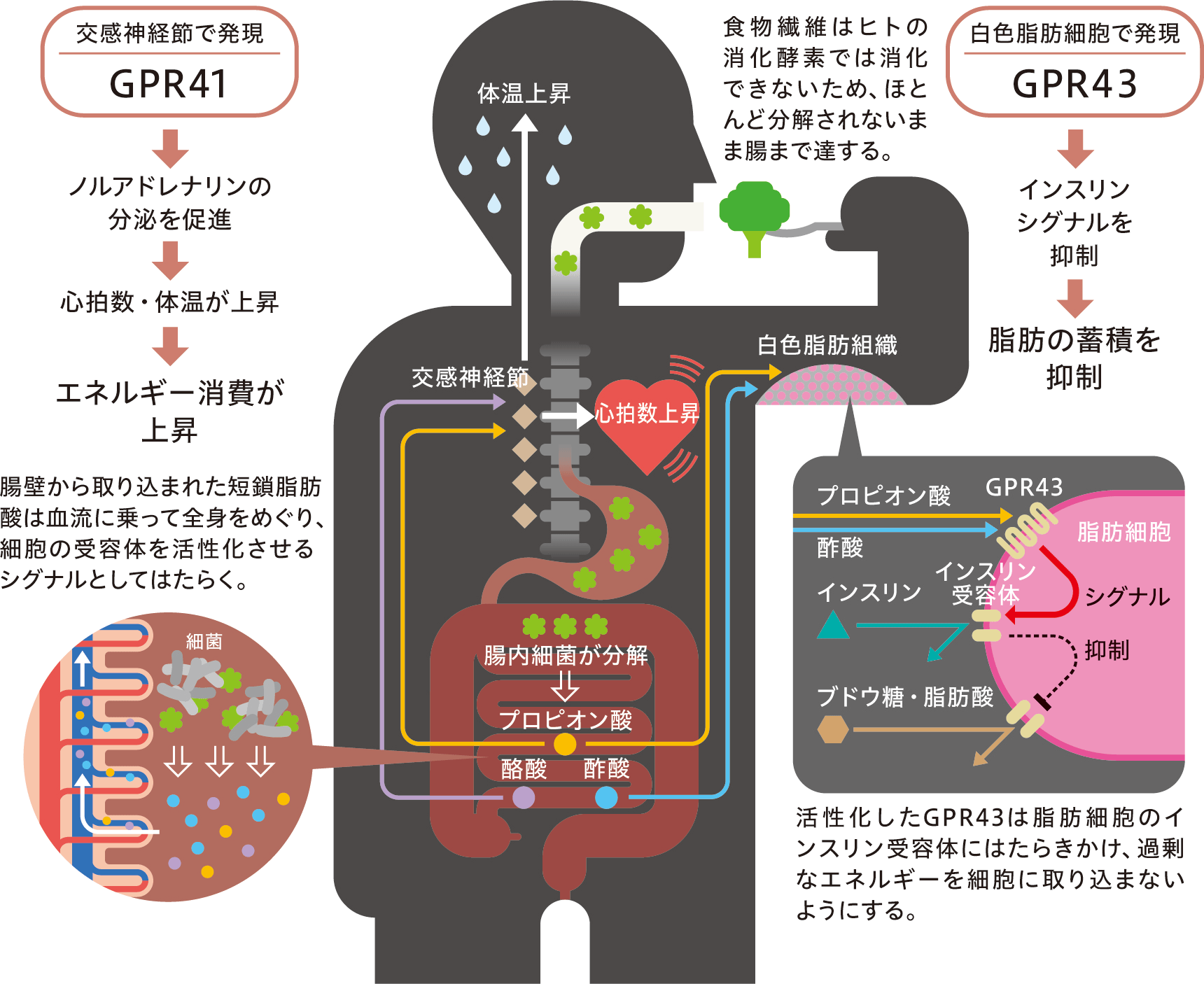
(図10) 短鎖脂肪酸によってはたらく、GPR41と43の肥満との関わり。

木村郁夫(きむら いくお)
2006年 京都大学大学院 薬学研究科 博士課程修了。 博士(薬学)。
2006年 千葉科学大学 薬学部 応用薬理学教室 助手・助教
2008年 京都大学大学院 薬学研究科 薬理ゲノミクス分野 助教
2011年 カリフォルニア大学サンディエゴ校 医学部 客員研究員 を経て、2013年より 東京農工大学大学院 農学研究院 応用生命化学専攻 テニュアトラック特任准教授














