RESEARCH
遺伝子の段階的な進化が生む
新しい植物の形
生きものは同じ遺伝子を異なる器官で使いまわすことがあります。被子植物のめしべではたらくCRC/DL遺伝子は、真正双子葉類では蜜腺で、単子葉類のイネでは中肋という葉の支持組織でもはたらき、それぞれの形づくりに寄与しています。この遺伝子は、どのような進化の過程を経て、多様な組織づくりに関わるようになったのでしょうか。
1.新しい形を生み出す遺伝子の進化
落葉の季節は楽しい。普段は高い位置にあって見ることのできない葉を、手にとって見ることができるからである。大きく迫力のある鋸歯を持つミズナラ、葉脈の二叉分岐が独自の雰囲気を持つイチョウ、公園や街路樹などでよく目にするケヤキなど、葉だけを見ても植物のかたちは美しく、そして多彩である。
これらの多様な陸上植物の祖先は、枝分かれした茎の先に胞子の詰まった生殖器官をもつ、極めて単純な構造であったと考えられている(図1)。それが長い年月を経て、光合成を行いやすい平面状の葉や、効率よく生殖できる花を獲得し、今日我々のよく知る植物のかたちとなった。生物の進化の歴史は、かたちの変化の歴史といえる。近年、生物のかたちの変化には、発生を司る遺伝子の進化が関係していることが明らかになってきた。
モデル植物として知られるシロイヌナズナの転写因子のひとつ、CRABS CLAW (CRC)遺伝子は、葉などの形成に関わるYABBY遺伝子群の中での遺伝子重複によって生じ、花の心皮(註1)と蜜腺(註2)の形づくりに関わっている。ところが、この遺伝子に相同なイネのDROOPING LEAF (DL)遺伝子は、シロイヌナズナで見られた心皮でのはたらきの他に、葉を直立させ、光を効率よく受けるのに役立つ中肋という支持組織の形づくりにも関わっていることがわかっている。ちなみにイネは蜜腺をもたない(図2)。さらに被子植物の中で最も起源が古い植物のひとつであるアンボレラでは、この遺伝子はめしべの心皮でしかはたらいていない(図3)。この遺伝子のもともとの機能は心皮でのはたらきであり、その後、シロイヌナズナでは昆虫を誘引する蜜腺、イネでは葉の強度を高める中肋の形づくりに関わるようになったのだろう。既にもっている遺伝子を別の場所に応用することで、新たな組織を獲得した興味深い例である。

左(図1)化石研究により示されている陸上植物の祖先の形態
初期の陸上植物のクックソニア。初めて枝分かれ構造を作った。
右(図2)イネの花
風媒花のため、昆虫を誘引する蜜腺は見られない。

(図3) 被子植物のさまざまな器官ではたらくCRC/DL遺伝子
遺伝子の進化というと、ゲノム上に存在する遺伝子のDNA配列が変化して、新たな機能を獲得するという印象が強いかもしれない。しかしこの例のように、はたらく時期や場所(発現パターン)を変化させることで、新しい器官や組織を獲得するやり方もある。そしてこれが形態の変化の重要な原動力のひとつになっているのである。
それでは、ここで注目したCRC/DL遺伝子はどのような過程を経て心皮、蜜腺、中肋と発現場所を多様に変化させてきたのだろうか。
(註1)心皮
めしべを構成する器官で、特殊に分化した葉とみることができる。将来、種子になる部分を包んでおり、種子を外界から保護する場である。受精後は果実の一部ないし全体になる。
(註2) 蜜腺
受粉を助ける昆虫を誘引するために蜜を分泌する腺。場所も形も様々であり、分泌細胞が集まっただけの組織レベルのものから、簡単な器官を構成しているものまである。
2.アスパラガスから見えてきたCRC/DL遺伝子のはたらきの変化の過程
単子葉類の中で、イネよりも古くに分岐したアスパラガス属は、真正双子葉類と単子葉類が分岐した後の初期の単子葉類の状態を保持していると考えられ、イネで見られた葉でのはたらきがどのような過程を経てきたのかを知るのに都合がよい植物群である(図4)。また、心皮の縫合線上にseptal nectaryと呼ばれるタイプの蜜腺を持つので、シロイヌナズナで報告された蜜腺でのはたらきも比べることができる。そこで私はアスパラガス属の一種であるクサナギカズラ(Asparagus asparagoides)でのCRC/DL遺伝子の発現を調べた。
まず、クサナギカズラのゲノム中にはCRC/DL遺伝子の相同遺伝子がひとつだけしか存在しないことを確認し、それをAaDLと名付けた。AaDL遺伝子の発現を調べると、心皮ではもちろん、イネと同様に葉でも発現していたが、その発現パターンは大きく異なっていた。

(図4)クサナギカズラの心皮と蜜腺(septal nectary)
三つの心皮が合着しためしべの内部に三つの蜜腺があり、心皮のすき間から蜜がしみ出ている。(左:草姿、右:心皮)
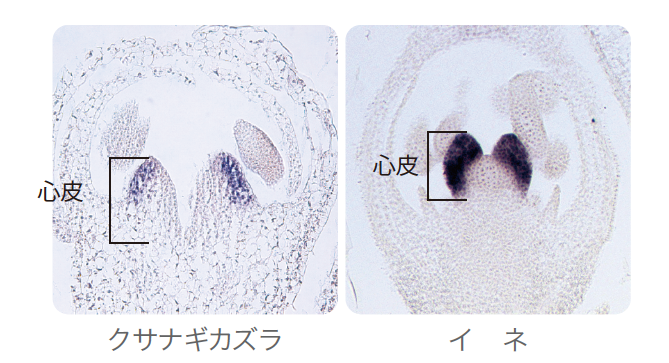
(図5)クサナギカズラ・イネの心皮でのCRC/DL遺伝子の発現領域の比較
青く染まった領域に注目。クサナギカズラでは心皮の一部のみで発現しているが、イネでは心皮全体で発現している。
まず心皮を見てみると、アンボレラとシロイヌナズナではこの遺伝子は心皮原基の背軸側(裏側)でしか発現していないのに対し、イネでは心皮の原基全体で発現している(図5右)。また、シロイヌナズナではこの遺伝子を欠いても心皮の形に異常をきたす程度に留まるが、イネでは心皮自体が作られなくなる。このことからCRC/DL遺伝子は、植物によって心皮の形成に寄与する度合が異なることがわかる。クサナギカズラはイネと同じ単子葉類でありながら、心皮でのAaDLの発現パターンはイネではなく、アンボレラやシロイヌナズナと同じであった(図5左)。このことから考えて、イネのように発現領域が心皮全体に拡大し、器官の運命決定まで担うようになったのは、単子葉類の系統でアスパラガス属の系統が分岐した後の出来事なのだろう。
次にイネに特徴的な葉でのはたらきはどうだろう。イネのDL遺伝子は、葉の発生初期には中央の未分化な葉肉細胞で発現し、その後、維管束以外の中央の葉肉細胞で発現する(図6左)。この未分化な細胞での発現が、葉の中央部の葉肉細胞の増殖を促し、中肋をつくると考えられている。一方、中肋のような構造を持たないクサナギカズラの葉では、師部細胞でしか発現していなかった(図6右)。葉の中央部での発現は、おそらく初期の単子葉類からアスパラガス属が分岐する以前に獲得されていたと考えられる。また、CRC/DL遺伝子の進化において、イネで見られる新しい機能の獲得は、発現領域の一度の変化でもたらされたものではなく、単子葉類の系統の中で発現領域が段階的に変化することでもたらされたのだと考えられる。
最後に蜜腺を見ると、クサナギカズラの蜜腺ではAaDLは発現していなかった。シロイヌナズナとは異なり、クサナギカズラの蜜腺の形づくりにCRC/DL遺伝子は関与していないようだ。蜜腺は、その形態、位置などが植物によって様々であり、それぞれの系統ごとに由来も異なる。蜜腺をつくる際に利用した遺伝子は常に同じではなく、その都度、都合のよいものが選ばれたことが伺える。

(図6)中肋の形成に至るCRC/DL遺伝子の発現場所の変化
3.CRC/DL遺伝子が様々な場所ではたらけるわけ
CRC/DL遺伝子は被子植物の進化の初期段階において、心皮の形づくりに関わっていた。その後に単子葉類に至る系統が分岐し、その中で心皮以外に葉でのはたらきを獲得し、さらにイネ科に至るまでの間に、葉の中肋を形づくるようになったと考えられる。蜜腺でのはたらきは、真正双子葉類の系統が他と分岐した後に獲得したのだろう(図7)。
それでは、CRC/DL遺伝子が心皮、葉の中肋、そして蜜腺という様々な場所ではたらく背景には何があるのだろうか。CRC/DL遺伝子は特定の細胞群の増殖を促進するはたらきを司ることがわかっており、私はこの機能が重要であると考えている。蜜腺や中肋は、それぞれの系統群で新たに獲得した構造である。新たな構造を生み出すためには、新たな部位で組織を肥大させる必要があり、CRC/DL遺伝子がもつ細胞群の増殖を促進するはたらきはそれにうってつけだったのだろう。CRC/DL遺伝子の発現領域の変化をもたらした分子機構を明らかにすることが今後の課題である。
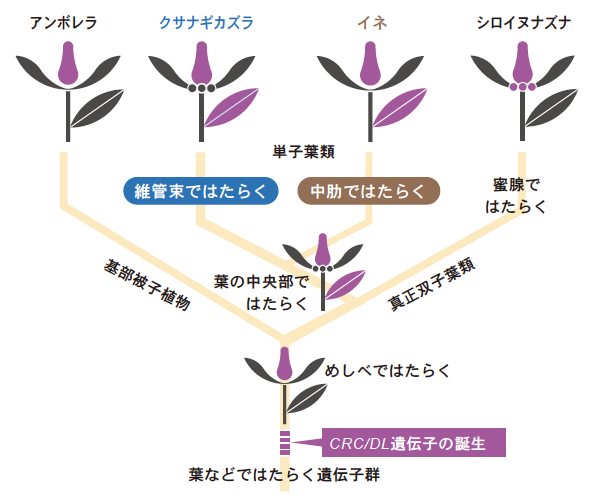
(図7) 段階的に発現場所を変えてきたCRC/DL遺伝子
4.植物の柔軟性
今回、私はCRC/DL遺伝子の心皮、蜜腺、葉での発現パターンを系統間で比較し、形の進化に伴う遺伝子のはたらきの変化の過程の一端を明らかにした。元々は心皮ではたらいていた遺伝子を、花の蜜腺や葉の中肋に応用した植物の柔軟性には改めて驚かされ、その過程は興味深いものだった。クサナギカズラは今回示した以外にも、側枝の位置に葉状の器官である擬葉(仮葉枝)をもつなど植物の形づくりの進化を探るうえで非常に興味深い特徴をもつ(図8)。私は、擬葉は枝の変形であると考え、これまでに葉の形づくりに関わる遺伝子が擬葉で発現していることを明らかにしてきた。アスパラガスが枝を葉のように変化させたのだとすると、これもまた驚くべきことだ。ここからも生物は既存の遺伝子を活用することで、様々な環境に柔軟に対応していることがわかる。
これまでの研究では、ごく少数のモデル生物の比較によって生物の共通性を掘り下げてきた。しかし近年、研究者が自身の興味をもつ事象を解き明かすために、さまざまな生物を対象にして研究できる環境が整ってきた。このようにして共通性と独自性を追ってこそ、生物の多様性が理解できると期待している。

(図8) クサナギカズラの茎頂部と擬葉の外形
クサナギカズラでは、枝が出る位置に葉状の擬葉が発生する。
中山北斗(なかやま・ほくと)
日本学術振興会特別研究員。2010年東京大学大学院理学系研究科博士課程修了。理学博士。現在、東京大学大学院理学系研究科発生進化研究室にて、植物の形づくりやその進化過程に関わる分子メカニズムを研究している。
.jpg)
.jpg)











