RESEARCH
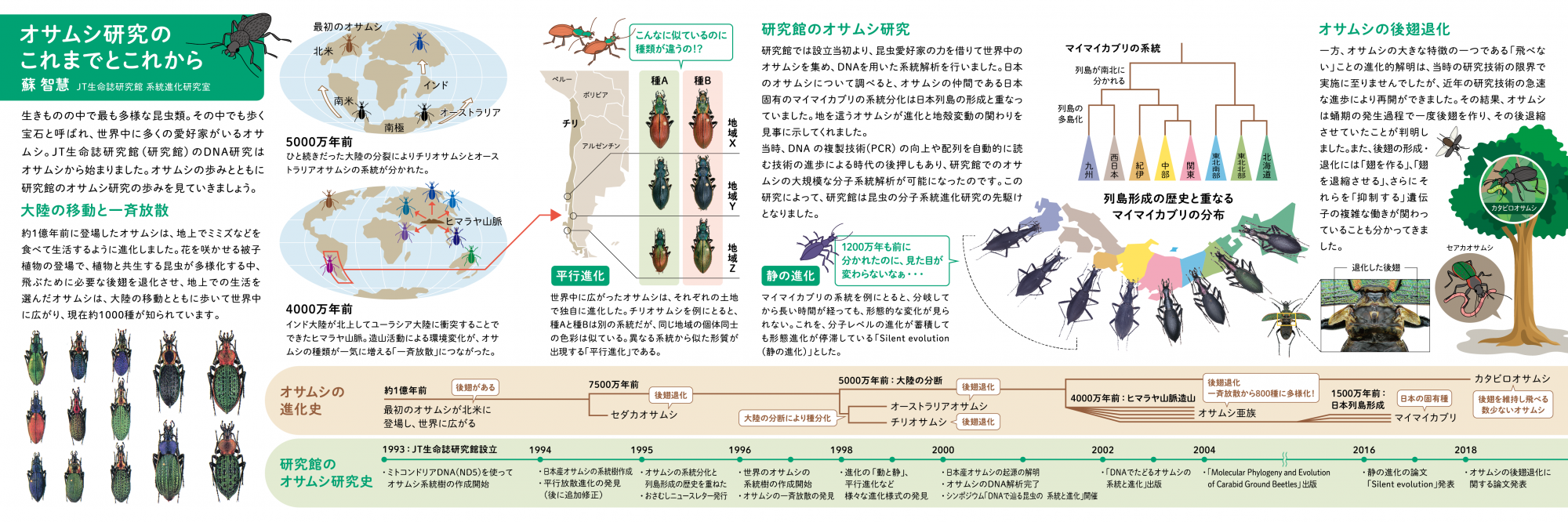
オサムシ研究のこれまでとこれから
JT生命誌研究館では、開館当初からオサムシの遺伝子解析研究を行なってきた。野外の昆虫を対象とした世界的なDNA調査は、日本の昆虫の分子系統進化研究の先駆けとなり、多くの成果を生んだ。私たちのオサムシ研究のこれまでとこれからの展望を紹介する。
本記事のまとめです。上図を右に移動してみてください。
1.研究館のオサムシ研究
生命誌研究館(BRH)の設立当初より、大澤省三博士(BRH初代顧問)が中心となって始めたオサムシの分子系統解析研究プロジェクト。当時はオサムシのような自然中の昆虫のDNAを用いた解析はまだ行われておらず、大きなチャレンジだった。半年後の1994年4月に私が研究チームに加わり、大澤先生と二人三脚で研究を進めた。
研究の成否を左右するのは研究資料の入手だ。頼りになったのは大澤先生の「虫屋」の人脈だった。まずは近畿のオサムシ研究グループの方々がオサムシ採集の協力を約束してくれた。さらに日本全国のアマチュアの昆虫愛好家約50名へ材料採集の依頼状を送ったところ、次々とアルコール漬けのオサムシ標本が送られてきて、嬉しい悲鳴をあげた。これらの方々のご厚意に報いるために、「おさむしニュースレター」を1995年から1999年(20号)まで発行し、新しい研究結果を逐次共有した。こうしてBRHのオサムシ研究は、プロとアマチュアが共同で研究を進め、発見の喜びを分かち合う見事なモデルケースとなったのである。
1996年にフランスの大学、1997年に中国科学院、1998年には韓国の大学との共同研究を開始し、BRHのオサムシ研究の輪は着実に世界へと広がっていった。1980年代後半からPCR(DNA増幅技術)、1990年代初めからはDNA塩基配列の自動読み取り技術(オートシーケンサー)が登場し、DNA塩基配列の解析が容易になった時代の後押しもあり、我々のプロジェクトは、当時最新の解析技術を駆使した研究として時代の先端を走っていた。その技術を学ぶために、全国から学生や研究者、虫の愛好家らがBRHに集まった。BRHのオサムシ研究が、日本の昆虫分子系統進化研究の先駆けとなり、その基礎を作ったことは間違いないだろう。
オサムシという昆虫がどのような経緯で多様化し、世界中で美しい種を生み出したのかの一端が見えてきた。またオサムシは、進化という現象について様々な興味深い様式を見せてくれた。開館から続いてきた研究から見えてきたことを、30年の節目に振り返ってみたい。そしてオサムシ研究の今後の展望について紹介する。
2.大陸の移動と共に広がったオサムシ
いわゆるオサムシは、分類学的には甲虫目(=鞘翅目)、オサムシ科、オサムシ亜科に属する昆虫で、世界各地におよそ1000種存在する(図1)。
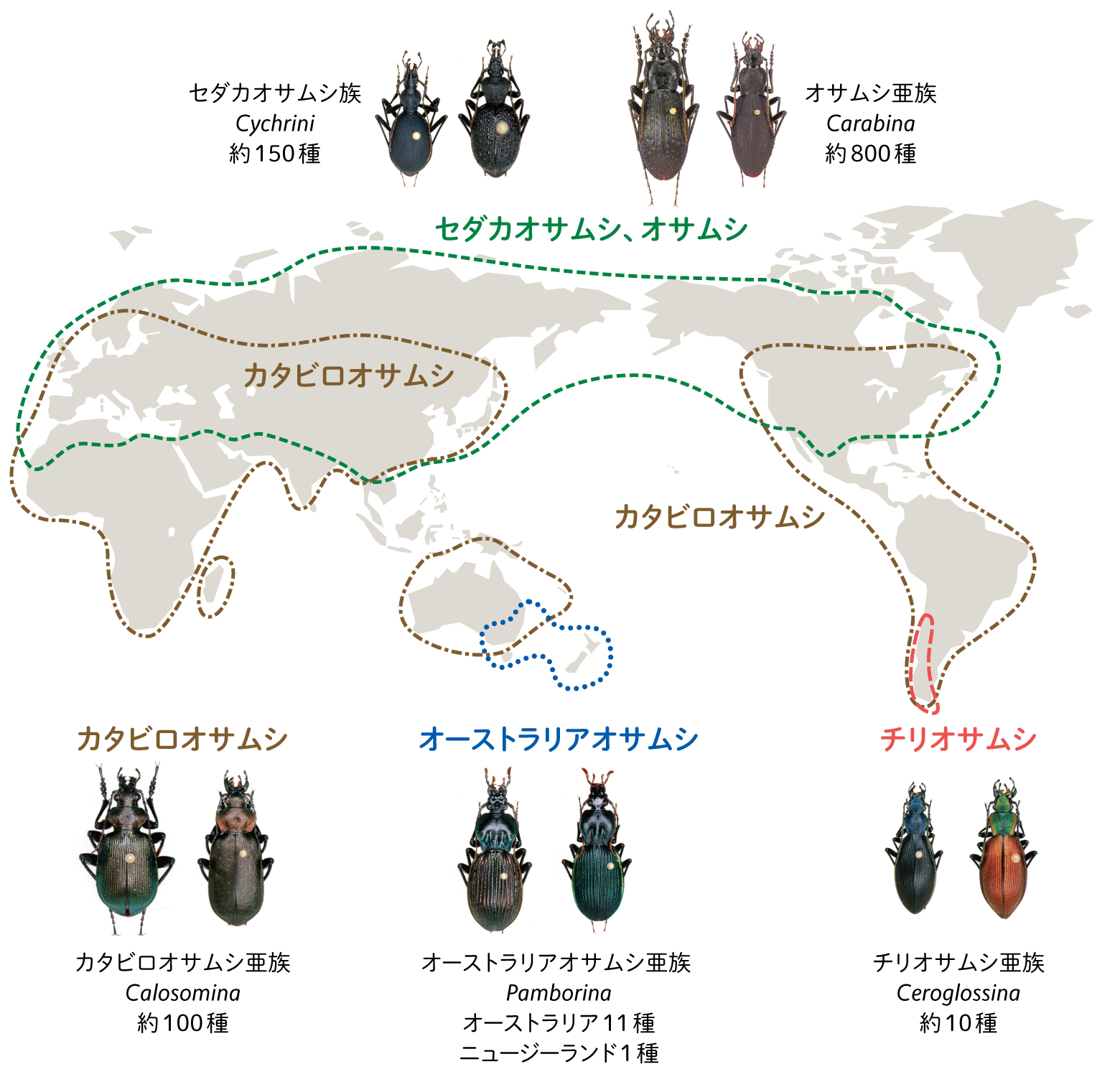
(図1) 世界のオサムシの分布
オサムシグループは、主に北半球に分布するセダカオサムシ族とオサムシ亜族、オーストラリアの南部とニュージーランドに分布するオーストラリアオサムシ亜族、南米チリとアルゼンチンの一部に分布するチリオサムシ亜族、そして北半球から南半球にかけて最も広く分布するカタビロオサムシ亜族に分けられる。種の数はオサムシ亜族が最も多い。
オサムシは、カブトムシなど他の甲虫類と同様、堅い前翅が体を保護する役割を担い、薄い後翅が飛ぶための役割を担う(図2)。しかし、オサムシは一部を除く全ての種が、飛ぶための後翅または飛翔筋、あるいはその両方が退化して飛べなくなっている。飛べないオサムシの中には美しく輝く前翅をもつものが多いため「歩く宝石」と呼ばれており(図3)、ヨーロッパをはじめ世界中に多くの愛好者がいる。それは我々の研究の資料集めに大いに役に立った。

(図2) 飛べないオサムシ(左)と飛べるカブトムシ(右)

(図3) 様々な世界のオサムシ
美しい前翅をもつ種が多く「歩く宝石」と呼ばれる。
我々は世界中のオサムシのDNAを解析し、系統関係と分岐年代の推定を試みた。当時、オサムシのような小さい分類群の分子系統解析には、核DNAよりも進化速度の速いミトコンドリアDNAが適当だと考え、ND5遺伝子を使用することに決めた。乾燥標本は遺伝子の増幅が全くうまくいかなかったため、協力者からの新鮮な標本が成果につながった(Osawa, Su & Imura, 2002)。こうして、オサムシの起源と、オサムシが世界中に分布するようになった経緯を以下のように推定することができた。
オサムシの祖先は、オサムシ亜科の外群となるゴミムシ、または現在特定できないオサムシ科の一種から、おおよそ1億~9000万年前に、現在の北米かそれに近いローラシア大陸で登場したようだ(図4上)。そして8000〜7000万年前、セダカオサムシ族が共通祖先から最初に分岐したとみられる。
その後、北米から南回りで、当時地続きだった南米・南極・オーストラリアへ広がったグループは、チリオサムシ亜族とオーストラリアオサムシ亜族に分かれた。2つの亜族が分岐したと推定される6000〜5000万年前は、南米大陸・南極大陸・オーストラリア大陸が分裂した年代とほぼ一致している(図4中)。つまりオーストラリアオサムシ亜族とチリオサムシ亜族は、これらの大陸の分断によって集団が隔離されたことによって成立したのだろう。(南極にいたオサムシは、南極大陸がその後南に移動したことで、寒さのために絶滅したとみられる)
一方、起源地である北米から北回りで、北極圏を経由し地続きだったユーラシアに広がったグループは、約5200万年前にオサムシ亜族とカタビロオサムシ亜族に分かれた。その後、4000〜3000万年前になると、2つの亜族の中で急速な種分化が始まる。このように短期間で様々な種が一斉に分化することを「一斉放散」と呼ぶ。4000万年前は、インド亜大陸がユーラシア大陸に衝突し、ヒマラヤ山脈やチベット高原が隆起した時代に相当する(図4下)。造山活動がオサムシの生息環境に大きな変化をもたらし、一斉放散の引き金になったと推測される。こうして種を増やした北半球のオサムシは、ユーラシア大陸全体に広がったのである。
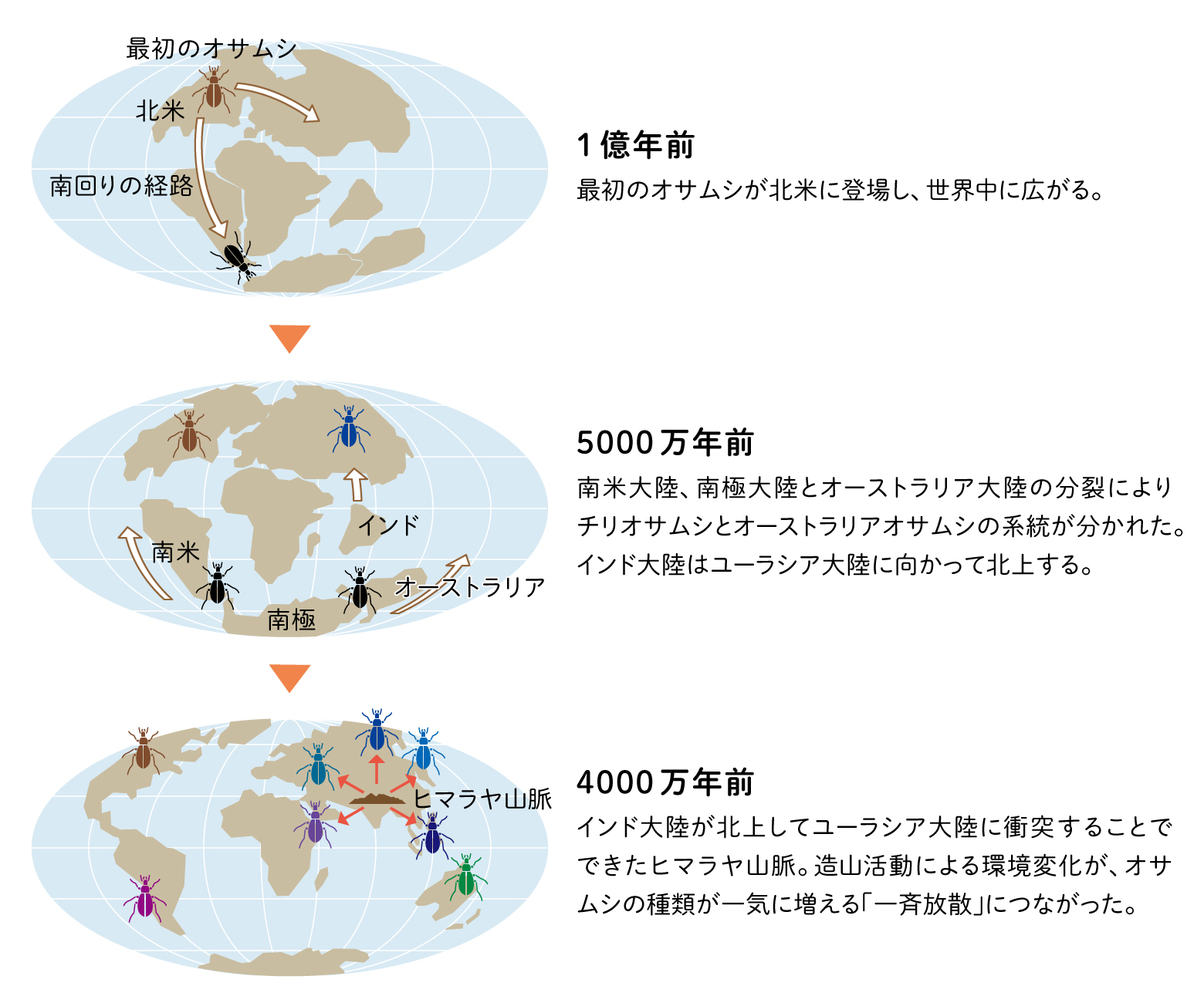
(図4) 地球上のオサムシの拡散・分化の歴史
3.日本列島の歴史を語るマイマイカブリ
日本にいるオサムシは、これまで全てが氷期(約 200 万年前以後)に日本列島に進入し、多様化したと考えられていた。しかし我々の系統解析の結果は、列島への進入経路が大きく2つのグループに分けられることを示した。
第1グループは、オオオサムシ亜属とマイマイカブリに代表されるグループで、約 1500 万年前、日本列島が大陸から分離する際にそこに乗っていた祖先種が、列島内で分化したと考えられる。第2グループは、ユーラシア大陸から氷期に陸橋を渡ってサハリンや千島経由で北海道に入ったグループ(アカガネオサムシ、コブスジアカガネオサムシなど)と、朝鮮半島から対馬に入ったグループ(ツシマカブリモドキなど)である。これらのオサムシは、氷期の直前に大陸やサハリンの種から分岐し、日本各地で多様化したことが系統解析の結果から推測できた。
第1グループに属するマイマイカブリの仲間について、特に興味深いことがわかった。このグループは約1500万年前にまず東西の2系統に分かれ、その後約1000万年前になると、それぞれがさらに3つと5つの亜系統に分岐している。興味深いことに、これら亜系統の分布は列島内の特定の地域に限定されている(図5)。これは日本列島が現在の姿になるまでに、まず大陸から離れて2つの半島になり、それらがさらに8つの島になる様子を写していると考えられる。まさにマイマイカブリの進化は日本列島の成り立ちを語っているようなのだ。
ちなみに、我々の最近の追加解析の結果から、西系統には、糸魚川・静岡構造線沿いにもう一つの亜系統が存在することが判明した。ここから逆算して、8つの島だった古日本列島は、実は9つの島だったのではないかとも考えられるのだ。

(図5) マイマイカブリの系統・分布と日本列島の歴史
4.平行進化の発見
オサムシの起源とその分布圏の成立について述べてきたが、オサムシがどのような経緯で多様化したのか、その進化の様式を知ることも研究の目的の一つである。作成した分子系統樹の枝の先にオサムシの形態の情報をのせると、進化の様子が浮かんでくる。形態の似たもの同士が必ずしも同じグループになるとは限らないことや、また、分岐してからの時間と形態的な違いの大きさが比例するとは限らないことなど、興味深い様々な進化現象を発見した。詳細は「大澤・蘇・井村、2000」を参照していただきたい。ここでは、代表的な「平行進化」の例について述べる。
進化的に離れた系統の間に似た形態が現れる現象を、平行進化と呼んだり収斂と呼んだりする。平行進化と収斂の区別は必ずしも明確でなく、同じものだという人もいる。また形態が全体的に似る場合と部分的に似る場合もある。ここでは便宜的にそれらを区別せず、すべて「平行進化」とする。
これまで、日本固有のオオオサムシ亜属や、クロナガオサムシ亜属など多くのオサムシグループから形態の平行進化の現象を発見した。ここでは一目で分かる、もっとも顕著な例の一つを紹介する。コウガオサムシCathaicus brandtiとクギヌキオオズオサムシEupachys glyptopterusである(図6)。

(図6) クギヌキオサムシとコウガオサムシ
2種はともに真っ黒で分厚い体の巨頭化オサムシだが、これらをヨロイオサムシ群(美麗なオサムシが多いグループ)の他の種とともに系統樹に入れてみてびっくり。見た目がよく似たこの2種は姉妹関係にはなく、コウガオサムシは美麗種揃いのカブリモドキ類(Coptolabrus)と、クギヌキオオズオサムシはこれまた歩く宝石クビナガオサムシ類(Acoptolabrus)と親類なのである (図7;Su et al., 2001)。忍者顔まけの見事な変身で、離れた系統から似た形態が現れる顕著な平行進化である。

(図7) オサムシの分子系統樹から見えた平行進化
もう一つ、チリオサムシの見事な色彩の平行進化を紹介する。図8を見てほしい。この図では、縦に産地を異にする同一種を並べた。縦に並んだ種が同種とは驚きである。また、横に産地(地域1, 2, 3, 4)を同じくする別種を並べたが、色だけで判断すると横に並んだ種は同じ種としか思えない。
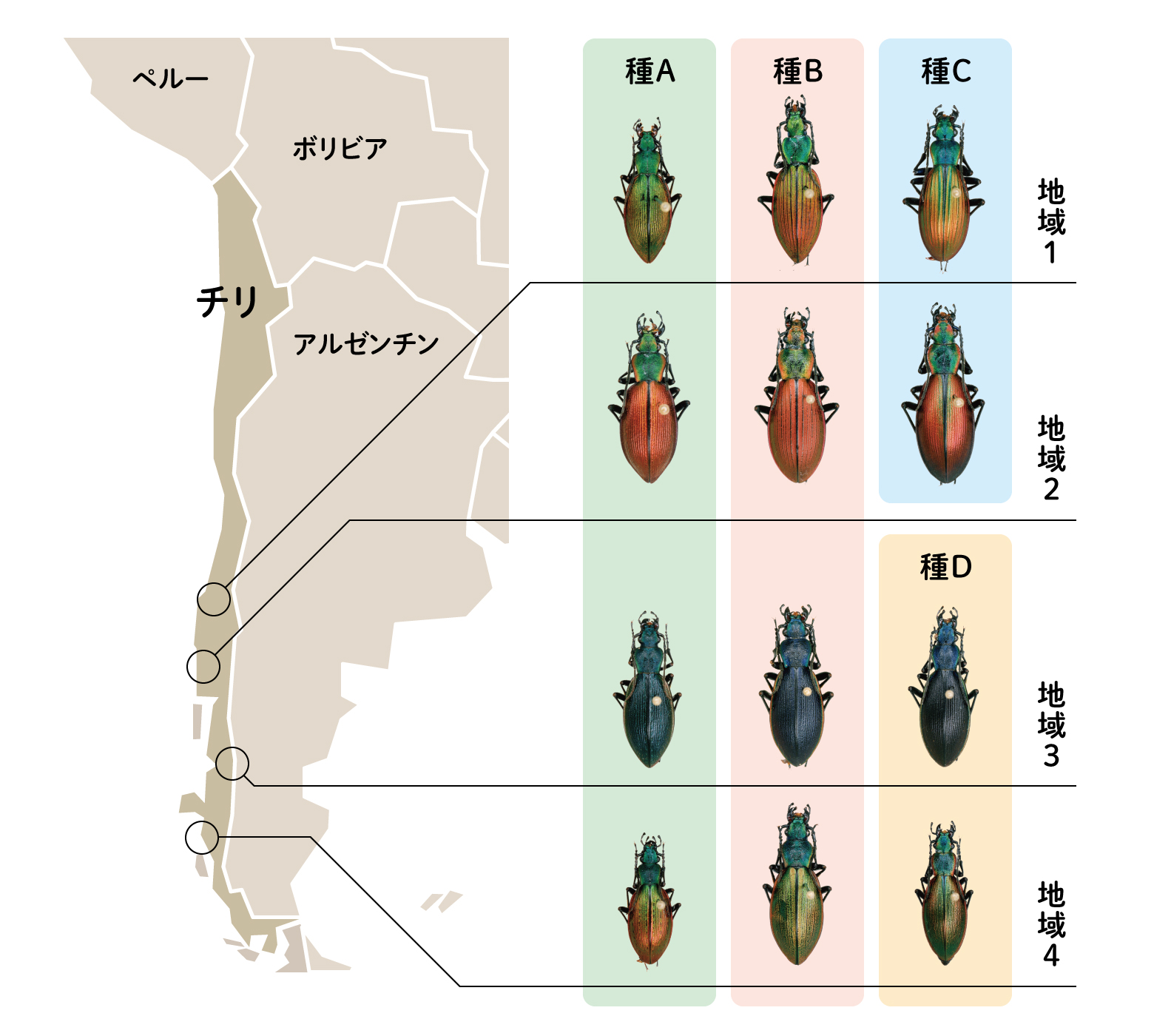
(図8) チリオサムシの色彩の平行進化
種A: Ceroglossus buqueti、 種B: C. chilensis、 種C: C. magellanicus、種D: C. darwini
これらは、当時学校教師であった柏井伸夫さんが採集されたチリオサムシである。色彩の多様性に惑わされてしまうが、分子系統や、詳細な形態的特徴を比較すると、縦列は同じ種と同定されるのだ。それでも“ほんとかいな”と思ってしまう。また興味深いのは、別種でも同じ所でとれるものは、色も似ているという規則性があることだ。このような色彩の平行進化がどうして起きたのかは、現状客観的証拠がないので推測になるが、一つは種分化前のチリオサムシに著しい色彩の多様性があり、それが種分化後に持ち越された結果、種の区別なく、それぞれの地域に特定の色のものが配分された可能性、もう一つは、初めはランダムにそれぞれの地域に配分された種が、地域の生息環境に対する適応進化の結果、色が分かれた可能性が考えられる。いずれの場合でも、系統樹からみると色彩の平行進化は比較的最近の出来事と推定される(Okamoto et al., 2001)。
5.静の進化と動の進化
分子系統樹は客観性が高いが、もう一つ重要な点は時間が系統樹に刻まれることである。分子進化は時間に比例して変異が蓄積される、分子時計の概念が導入されている。時間の要素が系統樹に入ると、系統樹から得られる進化的情報が俄然増える。形態の進化が起きていなくても、分子進化が進んでいる(時間が進んでいる)場合や、逆に分子進化があまり進んでいなくても(時間があまり経っていなくても)、大きな形態の進化が生じたことも分かる。
世界中のオサムシの分子系統樹を描き、その枝の末端に位置するオサムシを見渡してみると、顕著な形態変化が生じている2種がごく最近に分岐していたり、逆に形態の違いが全く見られない2種がかなり昔に分岐しているなどの例を多数発見した。前者は形態進化が短期間で起きた証拠であり、後者は形態進化が長期間に渡り停滞している証拠となる。我々は前者を「動の進化」、後者を「静の進化」とした(Su et al., 2001; 大澤・蘇・井村,2002;Osawa et al., 2016)。当然、「動の進化」と「静の進化」の時間は、相対的であり、絶対的な時間の基準は決められない。下記いくつかの例を紹介する。
「動の進化」の好例は、セアカオサムシ属 Hemicarabusの3種である。ヨーロッパセアカオサムシ、マックレイセアカオサムシ、セアカオサムシは一見して区別出来るほどの形態差があるが、これら3種の分岐はおよそ500万年前と、種分化としては比較的最近の出来事である(図9)。
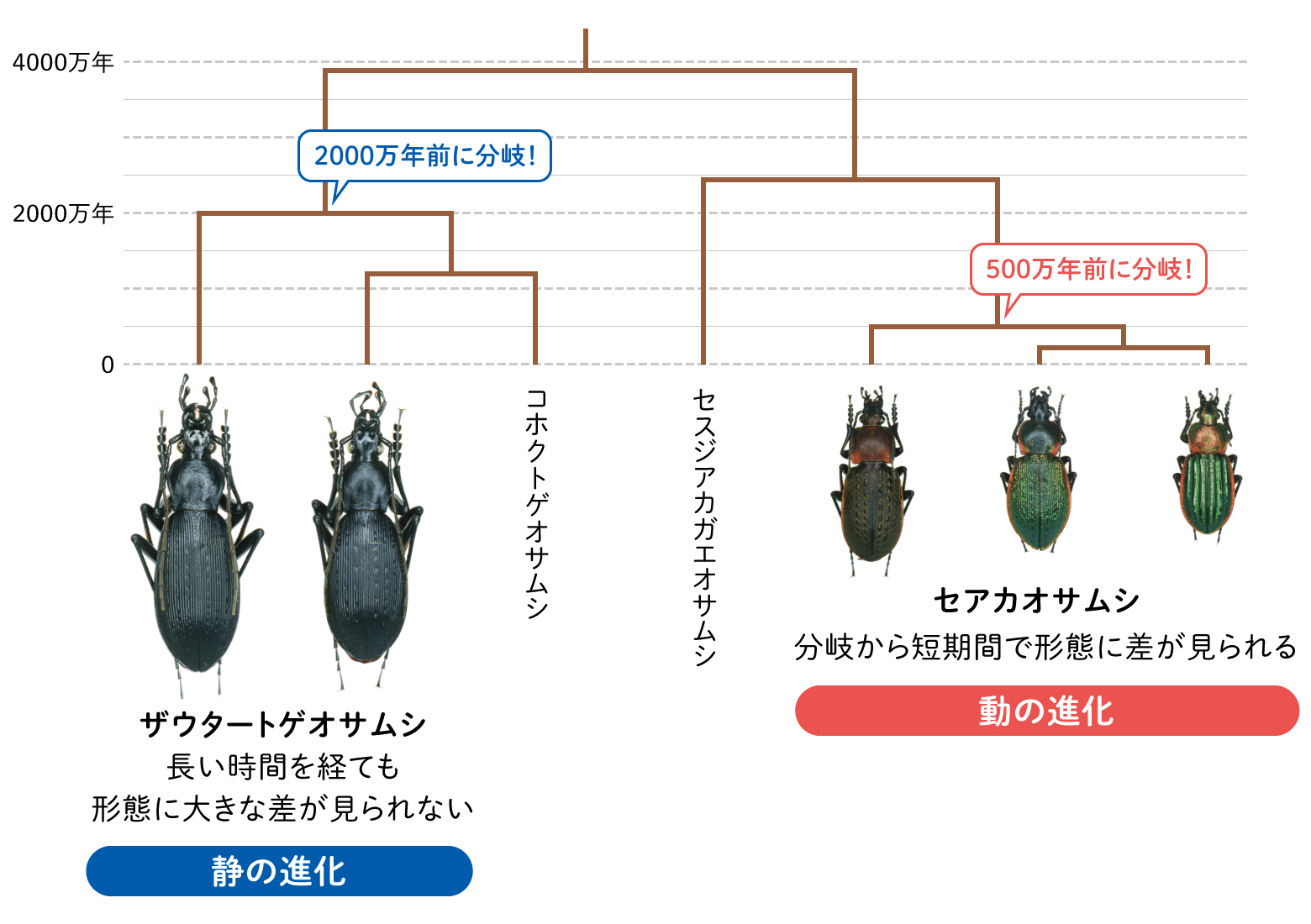
(図9) オサムシの分子系統樹から見えてきた「静の進化」と「動の進化」
「静の進化」については図9にも示したザウタートゲオサムシが好例であろう。2000万年以上前に分岐した三つの系統の間で、形態の違いが全く見られない。つまり、形態進化はおよそ2000万年にわたって停滞している。
日本国内の例を挙げると、日本固有種のアキタクロナガオサムシの進化は「動の進化」である。このオサムシはマークオサムシと近縁関係にあるが、マークオサムシから日本で約700万年前に著しい形態変化をともなって分岐している。その他にも、系統解析の結果から、国内のオサムシで、恐らく食性の適応進化によって巨頭化や狭頭化という進化が色々な分類群と地域において短期間で起きていることがわかっている。他にも、まだまだ「動の進化」「静の進化」の例をあげることができるが、ここでは割愛する。詳細は大澤・蘇・井村,2002とOsawa et al., 2016を参照していただきたい。
形態の変わらない「静の進化」がどうして生じているのかについては、恐らくそれらのオサムシの形態形成に必要不可欠な遺伝子群に変異が起きていないということになろう。もっと正確にいうと、そのような変異が起きると、適応度が下がるために淘汰されるとみられる。つまりそれらの形態は、長期に渡りもっともその生息環境に適応しており、安定維持されてきたということだ。勿論、形態や機能に変化をもたらさない変異は、遺伝子やゲノム上に蓄積され続けているはずである。
6.オサムシの後翅の退化
後翅が退化して飛べないことはオサムシの最も大きな特徴である。それによって地理的隔離が生じやすく、種分化や種の多様化につながりやすい。オサムシを研究材料に選んだのも、そういった進化の格好の材料としての特徴を考慮したためであった。
しかし、オサムシはなぜ、どのようにして後翅を無くしたのだろうか。オサムシ研究プロジェクトが開始した当初より興味深い問いの一つとして考えていた。しかし、後翅の退化的進化に関する研究は、当時の研究技術の限界で実施に至らなかったが、近年の研究技術の急速な進歩によって再開することができた。
まずオサムシ亜科内の各分類群の後翅の形態を詳細に調べることが先決であろうと考え研究を始めた。その結果、セダカオサムシ族の現存種に後翅はなく、前翅のみの2枚翅甲虫であることが判明した。一方、チリオサムシ亜族とオーストラリアオサムシ亜族では、後翅が1mmほどの短い針状に退化し、ほとんど痕跡のようなものとなっている。また、オサムシ亜族では、針状に退化したものから正常な後翅に近いものまで、退化後翅の形態が多様である。正常な後翅を持ち飛べるカタビロオサムシ亜族に、後翅が退化したものがいることが確かであることも判明した(Imura et al., 2018)。
以上の結果から、オサムシの後翅の退化は各分類群で独立に生じたと考えるのが妥当であろう。つまり、オサムシの祖先は正常な後翅をもち飛ぶことができた。カタビロオサムシの系統は飛翔機能を維持している。飛ぶことができた祖先の系統から、セダカオサムシ族、オーストラリアオサムシ族、オサムシ亜族が順次分岐し、その後、後翅が遺伝的な要因によってそれぞれのグループで独立に退化した。その遺伝的要因を突き止めるために、本研究は正常な後翅を持つカタビロオサムシ1種と後翅の退化形態が異なる代表的なオサムシ3種を用いて、後翅退化の発生過程の解明と、飛翔機能喪失の遺伝基盤と分子機構の進化の解明を目指している。
まず、4種のオサムシの飼育法を確立し、後翅の発生過程を観察した(図10)。その結果、前蛹期の後翅原基について、サイズと形態に明白な違いは観察されなかったが、クロカタビロオサムシの正常後翅原基は翅脈がより明確であるように見えた。また、退化後翅原基の中でマイマイカブリは最も翅脈が不明瞭であった。一方、蛹期ではカタビロオサムシの後翅は発生の進行につれて、翅脈間の組織が形成され、膜の中で翅が発達するが、退化後翅は、蛹期0日目では翅組織が膜の中で一杯になっていたが、2日目では翅組織の退縮が観察された。その後、ヤコンオサムシとマイマイカブリはその退縮がどんどん進行し、特にマイマイカブリの後翅は退縮程度が激しく、4日目にはすでに痕跡のように縮まっていた。ヤコンオサムシは羽化する前に後翅がすでに針状になっていることも確認された。一方、アカガネオサムシ(北海道タイプ)は、蛹2日目に翅組織の退縮が観察されたが、その後、翅組織が形成され発達する様子も見えた。羽化後のアカガネオサムシの後翅を観察したところ、その後翅は翅前縁から半分程度の大きさで、翅脈と翅脈間の翅膜は正常にみえた。
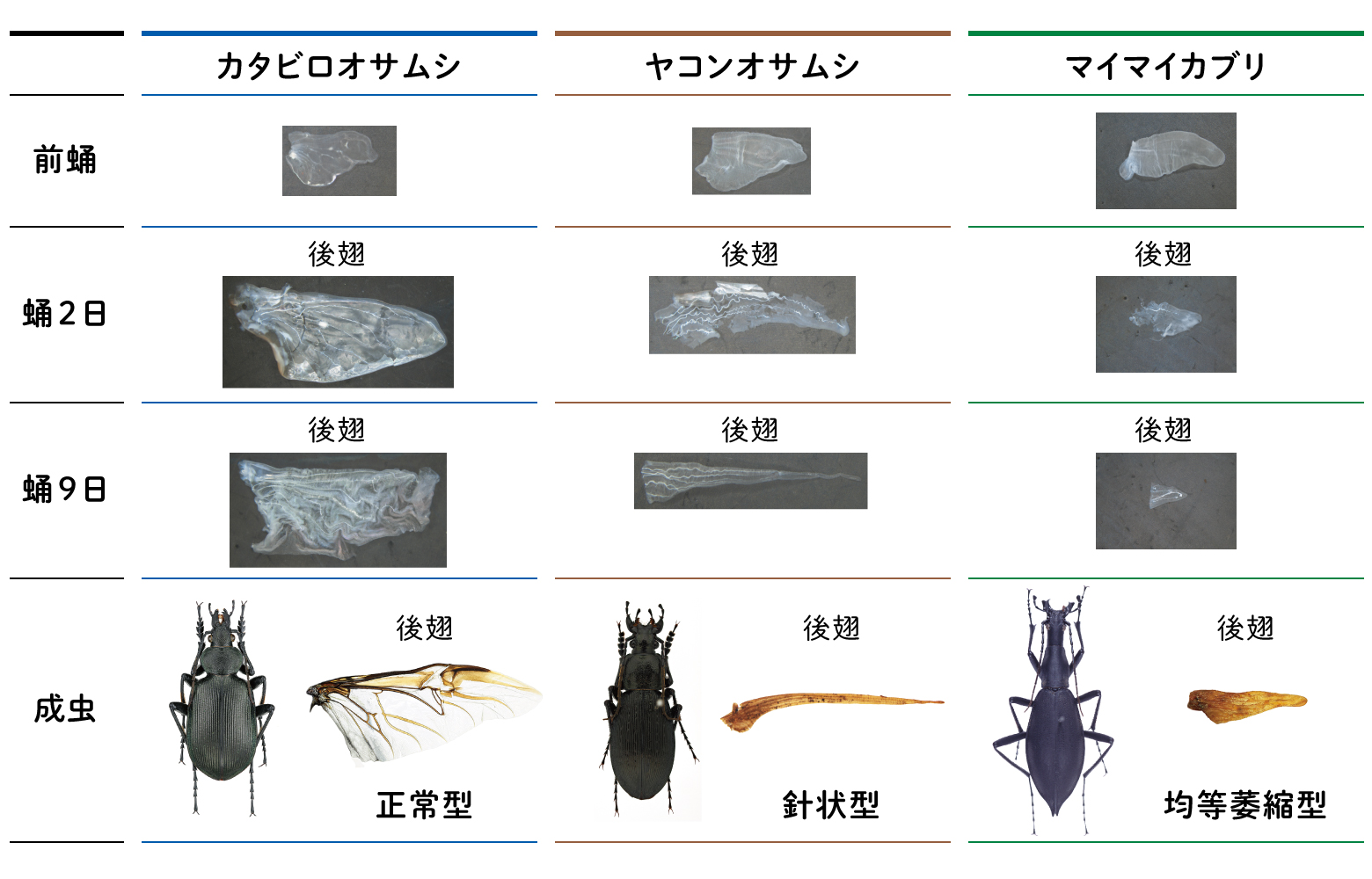
(図10) 様々なオサムシの後翅形成過程
以上の観察結果から、後翅の退化は蛹期の発生過程において、後翅の組織が正常に形成できないだけではなく、アポトーシスによる組織の退縮も関わっていることが明らかになった。また、RNA-seq解析による翅形成関連遺伝子の発現調査を行った結果、翅形成における翅脈間細胞分化に関わる遺伝子や翅の退縮を抑制する遺伝子が、正常後翅と比べ、退化後翅では発現が減少していることが判明した(図11)。今後はさらに候補遺伝子の特定を進めるとともに、候補遺伝子の機能検証も行う予定である。
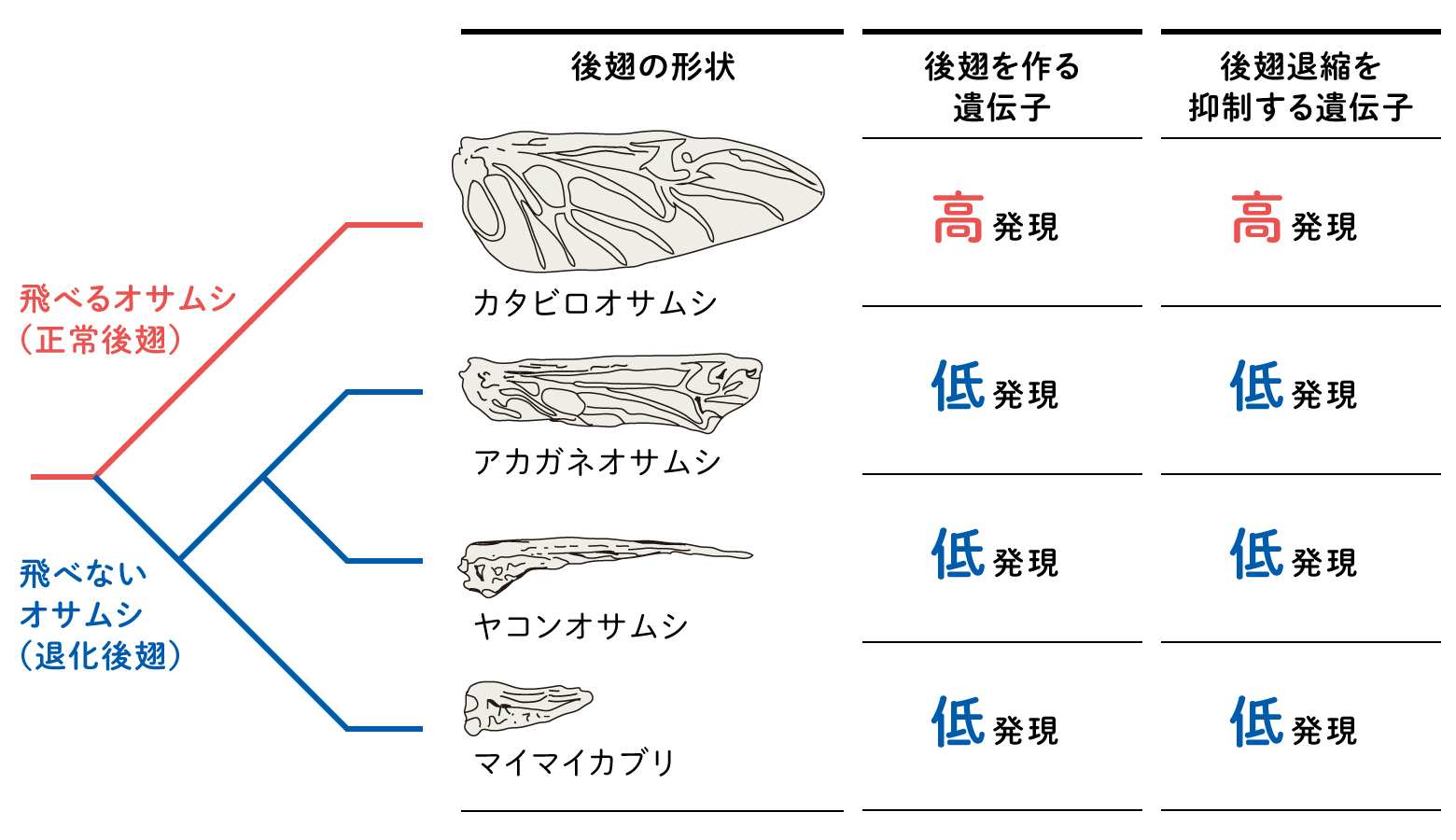
(図11) オサムシの後翅と系統関係
発生過程の観察や網羅的な遺伝子解析により、オサムシの後翅は祖先の段階で失われたのではなく、それぞれの系統で独立に退化したことがわかってきた。
7.研究に携わってくれた人々
冒頭で述べたとおり、BRHのオサムシ研究は、プロ・アマチュアも、国内外も問わず多くの人との共同作業により実現した。グループの一員であり日本のオサムシの生き字引でもある冨永修さん(当時大阪府職員)は、オサムシの産地を熟知しており、必要な資料を次々と届けてくれた。DNA解析の結果を伝えると、的確なコメントと共に不足の資料が調達されて返ってくる。このキャッチボールが「おさむしニュースレター」の発行に繋がった。1996年には、「世界のオサムシ大図鑑」を完成させたばかりの井村有希さんが加わり、研究は加速的に進展した。井村さんの本業は産婦人科医だが、ほぼプロのオサムシ研究者である。欧米のオサムシサンプルは、世界中にオサムシ研究者と愛好者の知人がいる井村さんが入手してくれた。1997年に東京大学で博士学位を取得した金衝坤さんがBRH奨励研究員として研究チームに加わり、DNA解析を担当した。その後、美しいチリオサムシに特別な興味をもつ柏井伸夫さん(当時・宝仙学園教師)と、北海道に分布する綺麗なオサムシが好きな岡本宗裕さん(現・京都大学霊長類研究所教授)も本格的に研究チームに加わった。柏井さんは毎年チリでチョウ採集の傍オサムシも採集し、次々とチリオサムシのサンプルを届けてくれた。岡本さんは、朝鮮半島、沿海州、北海道に分布するクビナガオサムシ類のDNA解析を担当した。オサムシについてここまでたくさんの研究成果を得ることができたのは、長期で深く研究に関わってくれた方々のお陰である。
.jpg)
蘇 智慧(そ・ちけい)
JT生命誌研究館 系統進化研究室 室長
カイコの休眠機構の研究で学位を取得しましたが、オサムシの魅力に惹かれ、進化の道へと進みました。1994年から現在に至るまで、ずっとJT生命誌研究館で研究生活を送ってきました。オサムシの系統と進化の研究から出発し、昆虫類をはじめとする節足動物の系統進化、イチジク属植物を始めとする生物の相互作用と種分化機構の研究を行っています。














