RESEARCH
イトヨの種分化を追って
性染色体の融合と種の分化
生きものは新しい種にどうやって分化していくのか。ダーウィンの時代から続く、進化生物学上の大きな問いの一つです。北野潤さんは、日本海と太平洋に住む2種類のイトヨに注目して、生殖隔離によって生じる種分化の解明に挑んできました。約200万年前の氷河期に始まったと考えられる日本のイトヨの種分化。果たして、どのようなゲノムの変化によってそれが実現されたのでしょうか。その遺伝基盤を明らかにする研究です。
1.生殖隔離による種分化
生きものは新しい種にどうやって分化していくのか。チャールズ・R・ダーウィンの時代から続く、進化生物学上の大きな問いの一つである。種分化の研究には、まず「種」の定義が必要である。エルンスト・W・マイアー(註1)は、種とは「お互いに交配しているグループで、他のグループとは生殖的に隔離されているグループ」と定義した。ここから、種分化の研究は生殖隔離が進行していく機構を明らかにすることだと言える。
実は、生殖隔離は連続的に進むことが多く、交雑が起きている場合も少なくない。そこで実際には種分化は、生殖隔離の程度が弱い集団から、完全に生殖隔離が完成している種まで、広い幅を持った連続性のある一連の現象として捉えることになる。
(註1) エルンスト・W・マイアー[Ernst Walter Mayr] (1904~2005年)
ドイツ生まれの進化生物学者であり、ハーバード大学名誉教授。主に鳥類の種分化を研究したが、生物学史にも造詣が深い。
2.種分化の研究モデルとしてのイトヨ
トゲウオ科(註2)に属するイトヨは、北半球の寒冷な地域の、淡水域から海水域にまで広く生息する体長3~8センチほどの魚である。この魚の興味深いところは、地域集団によって表現型が大きく異なり、地域ごとに程度の異なる生殖隔離が見られることである。例えば、カナダ西岸では湖底近くに住むイトヨと湖水面近くに住むイトヨが共存しているが、雑種は生態的適応度が低く、弱い生殖隔離が見られる。また、生涯を通じて河川の上流に住む小さいイトヨと、下流に住んで海に下る大きいイトヨの例では、中流域でこれらが接触しても、体の大きさが違うので交雑は稀にしかおきない。これらの集団は、いずれも人工授精では自由に交配でき、雑種が発生異常を示さないので、種分化のかなり初期段階にあると考えられる。
ところで、日本には日本海と太平洋のイトヨが生息しており、日本海イトヨは通常のイトヨ(G. aculeatus)とは遺伝的に大きく異なっている。そこで近年、G. nipponicusとして別種に区分された。約200万年前の氷河期に、日本海の地理的隔離が起き、そこから分化が始まったのだろう。また、北米の東海岸にも別種のイトヨ(G. wheatlandi)が生息しているが、通常のイトヨと交配しても稚魚は生存できず、完成された種分化の例といえる。このようにイトヨは、生殖隔離の弱い分化段階の集団から、隔離が完成された種までを含む種分化の連続体と捉えることができ、研究対象として興味深い。
(註2) トゲウオ科
少なくとも5属16種に分類されているが(魚のデータベース『FishBase』より)、1つの種の中にも多様な表現型を示す個体群がおり、さらに多くの種に分かれることが予想されている。
3.生殖隔離の遺伝基盤
種分化研究に私たちが選んだ日本海イトヨと太平洋イトヨは、北海道東部で生息域が重なる(図1)。ただし、雑種はほとんどいない。野外では生殖隔離が1つの機構で達成されることは稀であり、複数の機構の累積効果でもたらされる。日本海と太平洋のイトヨの場合も、複数の生殖隔離機構が同定できた(表1)。種分化の理解には、これら個々の機構が進化する遺伝基盤を明らかにすることが必須である。

(表1) 日本海と太平洋イトヨの間に存在する生殖隔離機構

(図1)地形の変化とイトヨの生息域
私たちはまず、行動隔離と雑種不妊に着目した。行動解析の結果、太平洋のメスは日本海のオスとはほとんど交配しないことがわかった。一つの要因は、日本海のオスが背中のトゲで強く求愛するため、太平洋のメスが逃げてしまうことにある(図2、動画1~3)。もう一つは、両種ともにメスは大きな体のオスを好むが、日本海のオスは太平洋のオスよりも体が小さいためである。一方、日本海のメスと太平洋のオスは交配するが、雑種のオスが不妊になることが明らかになった。

(図2) オスの求愛(左)とメスの逃避(右)
(動画1) 日本海オスから太平洋メスへの求愛
オスが求愛するとメスが逃げる
(動画2) 日本海のオスとメス同士の求愛
求愛を受入れる
(動画3) 太平洋のオスとメス同士の求愛
求愛を受入れる
これらの遺伝基盤を解明するために、連鎖解析(註3)を行い、ゲノム上の原因遺伝子の位置を調べた(図3)。その結果、求愛行動の違い、および求愛行動に利用するトゲの長さの違いを引き起こす原因遺伝子座が、染色体9番に局在していることが明らかになった。また、体の大きさの違いを引き起こす原因遺伝子座は、染色体19番に局在していた。さらに、雑種不妊は、日本海イトヨの染色体19番が太平洋イトヨの遺伝的背景と相互作用することによって起こることも明らかになった。イトヨでは染色体19番が性染色体であることが知られているので、これらの結果は、性染色体が種分化に重要であるとする理論と合致する。一方、染色体9番については予想外の結果が得られた。

(図3) 原因遺伝子の位置
(註3) 連鎖解析
DNAマーカーを利用して目的の遺伝子の位置を確定する解析法。本研究では、日本海と太平洋のイトヨ2種を交配して雑種1世代目を作成し、これを太平洋型のオスに戻し交配し、まず求愛行動と雑種不妊の表現型を解析した。その後、全ゲノムを網羅するSNP(一塩基多型)マーカーを利用して連鎖地図を作成し、ゲノム上のどの部位に原因遺伝子が存在するかを明らかにした。
4.日本海イトヨのネオ染色体
染色体の構造を比べたところ、日本海イトヨのオスの染色体9番がY染色体と融合していた(これを、新しく出来た染色体という意味でネオY染色体と呼ぶ)。残った9番と19番の相同染色体をそれぞれX1とX2染色体とすると、X1X2Y型の性決定機構となっているのである(図4)。太平洋イトヨでの常染色体である9番の染色体は、日本海イトヨの場合、メスとオスでは保持する数が異なることからX2染色体となる。そして、この領域に求愛行動の分化に関わる遺伝子座が局在していることが明らかになった。予想もしていなかった、生殖隔離の進化と性染色体の転換の関係が示唆されたのである。

(図4) 染色体の構造
この関係をゲノムレベルで検証するために、性染色体へと転換したネオY染色体に、常染色体とは異なる特徴があるか否かを調べた。全ゲノム配列を解読して種内で比較したところ、日本海イトヨのネオY染色体では、塩基配列レベルでX2染色体との分化が始まっており(図5)、アミノ酸置換も多く生じていることが明らかになった。加えて、ネオYとX2染色体の間で遺伝子発現量にも差があることが明らかになった。次に種間で、太平洋イトヨの常染色体(染色体9番)と日本海イトヨのX2染色体を比較したところ、アミノ酸置換と発現量の異なる遺伝子が多いことがわかった(図6)。また、種内では固定しているが、種間では異なる一塩基多型(註4)の数も増えていることが明らかになった。

(図5) 種内でのX2とネオYの分化
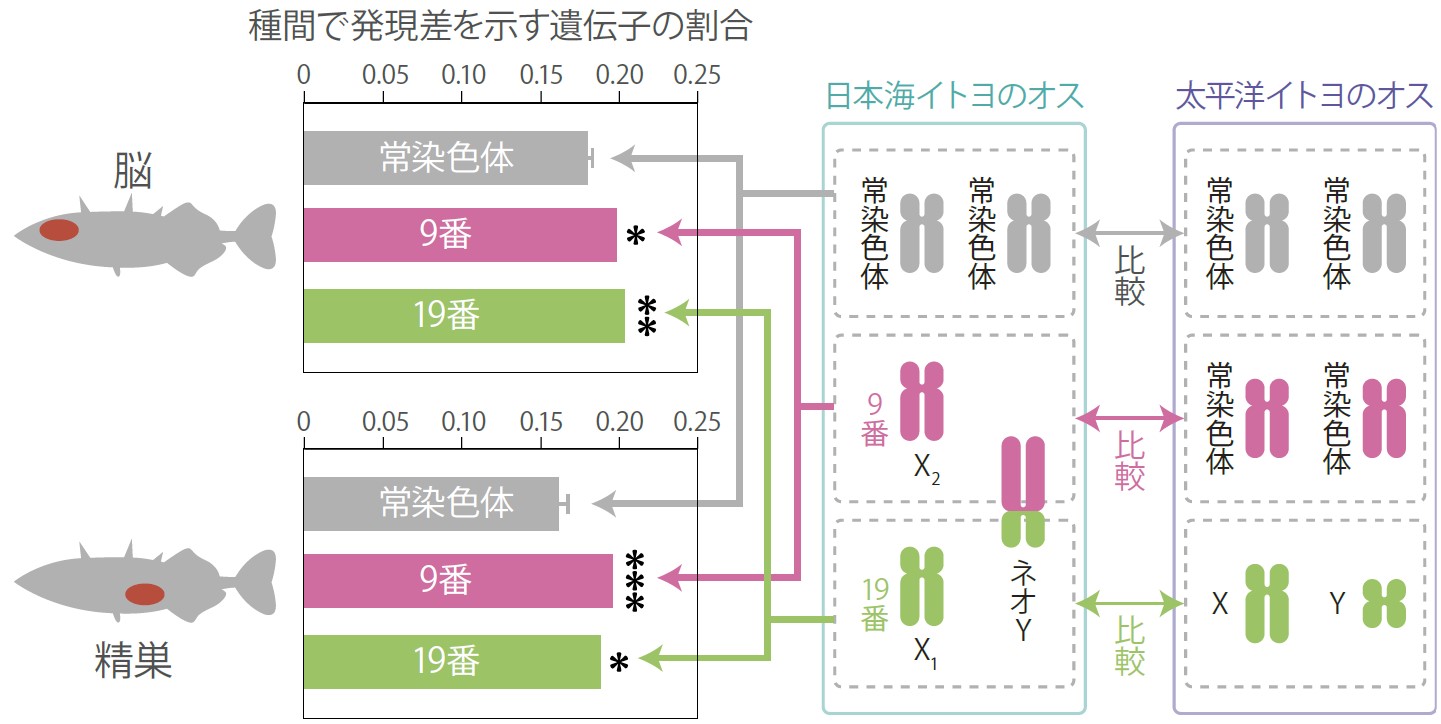
(図6) 種間での遺伝子発現の差
それぞれのグラフ中で、常染色体の値と比較した場合の有意差。
* P<0.05, ** P<0.01, *** P<0.001
(註4) 一塩基多型(SNP : Single nucleotide polymorphism)
個体間を比較した場合に、DNA塩基配列のひとつの塩基が他の塩基に置き換わっている多型。
5.野生生物の種分化機構の解明に向けて
北海道東部に生息する日本海と太平洋のイトヨについて、その種分化機構を解明する研究を始めて約10年になる。開始当初は全く予測していなかった、ネオ性染色体が種分化に貢献しているのではないかという仮説に行き当たり、その検証を続けている。思いがけない仮説の発見は、自然が我々の予測を超える何かを持っていると感じる瞬間であり、野外生物を対象とする研究の醍醐味と言える。
ここで紹介した全ゲノム解読の結果と連鎖解析の結果は、ネオ染色体への転換が、生殖隔離の進化を促進したと考える根拠となる。しかし、それに加えて染色体融合による組換え率の抑制など他の機構の貢献も考えられ、今後は個々の機構の貢献度を明らかにしていく必要がある。また、ネオ染色体の形成が種分化に貢献する例の普遍性を見ていく必要もある。性染色体の転換には、常染色体と性染色体の融合以外にも、性決定遺伝子が転移する場合もある。このような場合にも、ネオ染色体が種分化に関与しているのかを調べることは、私たちの仮説の一般性を検証する上で重要なことである。
TALEN(註5)やCRISPR/Casシステム(註6)などのゲノム編集技術によって、イトヨでも個別の遺伝子を破壊することが可能となった。この技術を使って、まず種分化や性的二型の進化に関わる候補遺伝子を特定し、その遺伝子を操作することで機能を確認したい。その遺伝子の振る舞いを野外で集団遺伝学的に解析し、野外生物の種分化の機構を知ることが私たちの研究の目的である。
(註5) TALEN [Transcription activator-like effector nuclease]
植物病原菌(Xanthomonas)に由来するDNA結合ドメインと、2本鎖DNAを切断できるヌクレアーゼドメインを融合したハイブリッド酵素。結合ドメイン中のRVD(Repeat variable diresidues)と呼ばれる可変領域を変えることによって、特定のDNA配列を認識・切断することができるゲノム編集技術。
(註6) CRISPR/Cas システム[Clustered regularly interspaced short palindromic repeat/ CRISPR-associated]
CRISPRは原核生物における獲得免疫系として働く座位であり、その繰り返し配列の近傍にはCas遺伝子群が存在する。本来、原核生物がCasタンパク質によって外来DNAを断片化し、CRISPR座位にそれを挿入することによって免疫記憶として残すための機能。任意のガイドRNAを用意することによって、特定のDNA配列を切断することができるゲノム編集技術。
図2、3:Kitano J, et al., Nature (2009) 461, 1079-1083より改変
図3、4、5、6:Yoshida K, et al., PLOS Genetics (2014) 10, e1004223より改変

北野 潤(きたの・じゅん)
2002年京都大学大学院医学研究科博士課程修了。医学博士。京都大学大学院生命科学研究科認知情報学講座研修員、同講座助手、米国フレッドハッチンソン癌研究所ポスドク研究員、東北大学大学院生命科学研究科生物多様性進化分野助教を経て、2011年より国立遺伝学研究所新分野創造センター生態遺伝学研究室特任准教授。














