



常時働く
子供の頃は普通の田舎の子供でした。中学校の音楽教師だった親父を筆頭に、年の離れた兄貴と姉貴も先生をしていたので、自分も教師になるのかなあとなんとなく思っていたのですが、それが本当に自分がなりたいものだという自信がなかったので、いろいろな大学をかけもち受験しました。しかも文系から理系まであらゆる学部を受けたのです。高校の教科書にDNAが登場し始めた頃で、それが基礎医学にもつながるかなという気持ちもあり、医学部も受けましたね。その頃は新潟に住んでいたのですが、どういうわけか北海道大学にとても憧れていて、文学部と医学部を受けました。結果はどちらもダメで、結局一浪して弘前大学理学部に入ったのです。あとは教員免許を取って高校の理科の先生にでもなるかと、僕も親もそう思っていました。
弘前大学は文理学部の改組で新しく理学部ができたばかりで、生物学科の学生はたった10人。少人数でわいわいやっているうちに生物学がだんだん面白くなってきて、先生になるにしても、大学院だけは行きたいと思い始めたのです。4年のときに入った斉藤和夫先生の研究室でバッタの染色体を観察したのですが、そこで卒業研究をした学生は、みんな染色体研究の大御所がいる北大の大学院に進学していたのです。僕もいよいよ北大だと思って希望したのですが、先生に「新潟大学なら入れる」と言われ、カエルの専門家岩澤久彰さんの研究室を紹介されました。
僕は岩澤さんについた最初の大学院生で、2人きりの研究室でした。毎日のスケジュールは、まず岩澤さんが7時前にやって来る。僕もなるべく早く来ようとするのですが、通学に時間がかかるので頑張っても8時になってしまいます。すると岩澤さんがお茶を淹れてくれて、いま生物学でどの先生がどんな仕事をされているかという話を30分くらいしてくれるのです。それが終わるとそれぞれ自分の仕事に入り、昼まで一言も交わさない。それを1年間続けているうちに、「研究者は常時働いているもの」という感覚が体にしみつきましたね。今でも取り柄はあまりないけれど、働くということだけはやり続けています。

11歳の頃。弟(左)と。

高校の修学旅行で。(右端:本人)
京大動物学教室へ
岩澤さんのもとでカエルの性分化に取り組み、その縁で博士課程はメダカの性分化を研究している東京大学動物学教室の江上信雄先生につくことになりました。メダカのオスとメスの外見上の差は成長してからでないと現れません。でも、生殖腺を見ると、孵化直後から生殖細胞数に違いがあって、卵をつくるか精子をつくるか区別できるんです。その時の僕の再発見です。当時メダカの性決定は性ホルモンで支配されているという説が有力だったので、孵化直後の早い時期の生殖腺にホルモン分泌細胞が存在しているかという問いを立て、切片を作って電子顕微鏡でくまなく探しましたが、1年近く観察しても見つかりませんでした。そこで、メダカの性分化はホルモン以外の仕組みで決まるのではないかという論文を出し、学位を取りました。次の行き先を探していたとき、京都大学の動物学教室で岡田節人先生の後任に入られた白上謙一先生が助手を探しておられたので応募し、これが僕と発生学の出会いになります。
東京大学でももちろん発生学の研究はあったのですが、京都に来てそのレベルの差に驚きました。東大では、フランス帰りの水野丈夫先生がニワトリの消化管をガラス皿に取り出して培養できるというのが最先端の仕事だったのですが、京大ではイギリス帰りの岡田先生が細胞1個1個を培養する技術を確立されていた。議論されている内容もまったく違っていて、南アフリカ出身の発生学者が複雑な体の形ができ上がる発生のモデルを幾何学的に説明したという世界の先端の話題がとりあげられていたのです。僕には全くの初耳です。白上先生も、最初は「君も知っているだろうけど」という前置きで話を始めていらしたのに、そのうち「君は知らないだろうけど」から始めるようになられて。多分呆れていらしたと思いますよ。そんな状態の中で、カエルの原腸が陥入して形態形成を始めるときに個々の細胞はどんな構造をとるか、電子顕微鏡で観察するという仕事を始めました。


Scientist Library:
生命誌30号
『ルイセンコの時代があった 生物学のイデオロギーの時代に』
岡田節人
ホヤとの出会い
白上先生は当時としては非常に珍しく、発生を動画で観察する重要性を知っていた方でした。顕微鏡を用いた微速度撮影装置を持っていて、ヒキガエルの卵が分裂していく様子をじっくり調べられた。細胞はぽこぽこと瞬間的に分裂するんじゃないんですね。分裂が波が伝わるようにじわーと広がっていく脈動運動という現象がある。これを突き止められたのです。1日かかる初期発生の様子を3分間くらいの動画にしたものを先生が自慢そうに見せてくれる。その後で、さあ自分の仕事だと一階の電子顕微鏡室にこもるんですが、僕の場合は何百枚も切片を観察してやっと1つの時期の1つの卵を見たことにしかならないわけです。なんというかそのギャップがものすごくてね。発生学って何だろうと考えると、今でもちゃんとわかっているわけではないんですが、時間という動的なものを入れない発生学はありえないとその時強く思った気持ちが今も続いていますね。
そこで、テーマを変え、受精卵が分裂して原腸が陥入するまでの時間がどうやって決まるかを調べようと考えました。しかしカエルは原腸が陥入するまでに14回分裂します。何分後の何回目の卵割でいくつの割球ができたという記述をしていくと、最終的には2の14乗ですから1万個以上の細胞を数えなければなりません。これは大変なので、もうちょっといい材料はないかなと思いましたが、ここではカエルの研究をするのが助手採用の条件だったのです。
ところが、僕が京大に来て1年も経たないうちに白上先生ががんで亡くなられ、助教授の方もその1年後に急逝され、助手1人がぽつんと残される事態になったのです。困りましたけど、こういう時は開き直って自分で考えるしかありません。そこで、カエルをやめて原腸陥入までの時間を見るのに都合の良い生きもの探しを始めました。白上先生の著書をあらためて読んでみると、やはり先生はよく勉強なさり、考えている方でした。カエルの原腸陥入と比較して、同じ脊索動物でもホヤは7回目の分裂後わずか120細胞ほどで原腸が形成されると書いてあったのです。これだと思いましたね。僕とホヤとの出会いです。

白上先生と。京都大学瀬戸臨海実験所の前で。
発生の時間を計る
当時日本ではホヤの初期発生の研究は行われていませんでした。ホヤには群体をつくる仲間があるのですが、そのホヤが2匹出会ったときに融合するかしないかは遺伝的に決まっているとか、あるいは単体のホヤは雌雄同体なのに自家受精しない仕組みがあるというような、自己認識の実験は盛んに行われていたのですが。僕は東北大学の臨海実験所にホヤの扱い方を習いに行き、東北地方を中心に分布するマボヤで発生研究を始めました。

左.成体のマボヤ。 右.研究棟の地下で飼育中のカタユウレイボヤ。
発生の一定時間後に原腸陥入という形態形成が始まるのは、細胞分裂の回数をもとにして時間を計っているからだと考えて研究を始めました。細胞が分裂するのに必要な要素としては、DNAの複製、核の分離、細胞質の分割の3つがある。どれが一番重要かをチェックしていこうと考えました。そこで先行研究を調べると、驚いたことにアメリカのウィタカーという研究者が、ホヤで面白い研究をしていたのです。薬剤で細胞質分裂を止めても、一定時間後に筋肉になるはずの割球にはちゃんと筋肉をつくるタンパク質が出てくるという報告です。発表されたばかりでした。彼は世界中に分布するカタユウレイボヤで実験していたので、あわててマボヤで追試すると、同じ結果が出ました。その薬剤処理では、核と細胞質の分裂が阻害され、DNA複製は行われています。当然、DNA複製を止めるとどうなるかが問題となるわけですが、ホヤの卵割は非常に速く進み普通のDNA複製阻害剤を用いてもなんの影響もないのです。ウィタカーも僕もここで足踏み状態が続きました。
ある時、東京大学農学部の池上晋先生が、菌から抽出したアフィディコリンという抗生物質がさまざまな生物のDNA複製を阻害する効果があるのだけれど、ためしに使ってみませんかと言って下さったのです。偶然の出会いでした。確かにアフィディコリンを使うとDNA複製がぴたっと止まる。ある回数以上DNA複製しないと、筋肉や消化管をつくる遺伝子がはたらいてこないという非常にきれいな結果が得られました。生物の発生を支配している時計の一つは、DNA複製の回数であるということが明らかにできたのです。一連の成果を『初期発生における細胞分化・形態形成開始の計時機構』という総説にまとめ、日本動物学雑誌に掲載しました。これが僕が若い頃の一番よい仕事だったと思います。
この総説を書いてから数年後、アフリカツメガエルで、12回の卵割が終わった胞胚期のところで割球の性質がいろいろ変わるという現象がわかってきて中期胞胚遷移と名付けられました。初期発生における時間の意味、時計の重要性が広く認識されるようになったのです。ただ、僕は、この先鞭をつけたことでよしとし、あえて競争に乗り込もうとは思いませんでした。実は、総説の最後を「機会を改めて、時間の流れを基にして初期発生を考えてみたい」と結んだのです。この時頭に描いていたのは、発生を、生きものの持つもう一つの時間である「進化」とつなげることだったのです。発生と進化の関係は多くの人が考えてきたことでしたが、当時は進化を研究する方法論はなかったので、実験発生学者として進化を扱うことはできませんでした。研究者の中でそれはテーマではないとされていたのです。僕も実際に進化に取り組んだのは、もう少し後のことになります。

東北大学浅虫臨海実験所での研究集会に来日したホヤ発生学の友人ウィリアム・ジェフェリーと。
学生が広げるホヤ研究の輪
ホヤは典型的なモザイク卵モザイク卵一部の割球を除去すると、それに対応して各器官が欠如した体をつくる卵。これに対し、残された割球で正常な体をつくることのできる卵を調節卵と呼ぶ。です。オタマジャクシ型の幼生をつくるのですが、幼生の細胞数が少なく、どの細胞がどの器官になるかを追いやすいという利点があります。まず研究室に来る学生のテーマは、発生学の基本である細胞分化と決め、細胞の系譜の追跡と分化決定因子(デターミナント)の探索を始めました。ここで最初によい仕事をしたのが、西田宏記さん(現大阪大学教授)。手先が器用だったんです。割球の1つ1つに目印をつけ、発生が進むとそれぞれがどんな細胞になるかを詳細に調べたのです。その結果、1世紀近く信じられていた細胞運命予定図に誤りがあることを発見しました。西田さんは独立してからもホヤ研究を続けて、筋肉をつくる割球のデターミナントが、卵形成のときに母親がつくりだすmRNAであることを突き止めるなど、次々よい仕事をして活躍しています。
西田さんに限らず、僕のところで学位を取った学生さんは、なぜかホヤを続けている人が多いですね。僕が頼んでいるわけじゃありません。大学などで職を得たのも本人の実力ですし。結果として、日本のホヤ研究の広がりに大きく貢献したことになっているわけで、ありがたいことです。
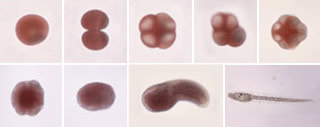
カタユウレイボヤの発生様式。わずか18時間で2,600程度の細胞からなるオタマジャクシ型幼生になる。(写真提供:西方敬人・甲南大学理工学部教授)
僕の方針として、学生さんが一度選んだテーマは、その人のものであり、その人が作り上げるものであるという考え方を徹底しました。だから、研究室を出ていっても続けたいという場合には、どんどんやりなさいと言って、僕は手を引くんです。僕は別のテーマを考えるわけです。自分ならホヤで面白いテーマをいくつでも見つけて成果を出せる自信があると、そう思い込んでいたからそうしたのです。でも、今考えてみると、もし同じテーマでやることにしていたら、むしろ負けるのは僕の方だったのではないかと最近思うようになりました。僕のところを出たホヤ研究者の活躍ぶりを見ていると、この世界で僕が生き残れているのは実は彼らとの競争を避けたおかげではないか。つくづくそう思います。

1991年、日本動物学会賞受賞記念パーティーにて。
発生と進化をつなぐ
1987年のある日、発生学のライバルである団まりなさん(元大阪市立大学教授、現階層生物学研究ラボ)が『個体発生と系統発生』と題する著作を送ってきてくれました。同封の手紙には、「自分は自分の考えを本にしてきちんと著した。これが本当の生物学者のやることである。おまえのようにくだらない論文を山ほど書いているのはさみしいことだ」とありました。早速読んでみると、序章に「比較発生学や、系統発生と個体発生の問題に興味を持つこと自体が、発生学者としての体質の古さを示すものとされるようになり、この問題は、ついには一種禁句のようになってしまうのである」とあったのです。僕は即座に「全くそうは思わない。問題の大きさに直面して筆を持てないでいるだけ」と反論のメモを書きました。団さんはこれまで行われた発生研究のデータを用いて、さまざまな動物の関係を見るという試みをされたわけだけれど、僕はまだ今の段階ではそれはできないと考えていました。興味はあるけれど、あまりにも大きなテーマで何をすればいいのかわからなかったのです。

団まりなの著書への反論メモ
ところがその翌年、主要な動物群でのリボソームRNAの配列を比較することで形態情報によらずに系統樹が描けるという論文が出ました。分子系統学の方法論を動物分類に応用した論文で、アメリカの研究者が発表したのです。これはすごい仕事だと思い、すぐに脊索動物の進化にかかわる分類問題の解決に乗り出しました。
ホヤと脊椎動物は脊索動物門の一員であり、ウニやギボシムシなどと同じ新口動物新口動物初期胚で形成された原口が、成体の口にならない動物の総称。後口動物ともいう。これに対し、原口がそのまま口となるものを旧口動物と呼ぶ。です。当時は、新口動物の仲間に他にもヤムシやハオリムシなど明瞭な脊索構造を持たないものがいくつも含まれていたので、このあたりの再検討から始めようと考え、これらの動物を含む分子系統を作りました。試みたのは和田洋君(現筑波大学教授)で、発生過程で脊索をつくる脊椎動物とホヤとナメクジウオとは、それ以外の動物群とははっきり分かれることが示せました。モデル生物以外で、日本の動物学に分子系統学を導入したのは彼が初めてでしょう。まだ修士の学生だった和田君が学会発表すると、僕のところに動物学の大家がわっと集まってきました。「今までの分類が覆る結果だが、あいつの言ってることは本当か」と口々に言うのです。僕としては脊索動物の起源が脊索の進化にあることがこれで確実になったと考えていました。ホヤの脊索こそ発生と進化をつなぐテーマであり、これで団さんに対抗できる「きちんとした研究」ができると興奮していました。
ウッズホール代理講師
分子生物学の技術を用いて発生と進化をつなぐ研究は、海外ではすでに始まっていました。僕は分子生物学の素人でしたが、偶然に海外でそういう研究をしている人々と知りあう機会が巡ってきました。アメリカのウッズホール海洋生物学研究所は、毎夏著名な研究者を講師に呼んで発生生物学のサマースクールを行っています。そこに講師として僕が呼ばれたのです。当時のサマースクールのまとめ役は、ホヤの研究者ウィタカーとウニの研究者エリック・デビッドソンだったのですが、ある時2人が仲たがいして(?)、ウィタカーのかわりに僕に白羽の矢が立ったというわけです。実はウィタカーはDNA複製停止の実験で僕に先を越されて以来、僕を目の敵にしていたのですが、その彼の後任に僕がなったのもなにかの因縁でしょう。
エリックはすでに分子生物学の大御所で、ウニの発生を遺伝子で追った研究論文をいくつも発表していました。他人の研究に容赦ない批評を浴びせる厳しい人でしたが、彼から学ぶことは多かったですね。もう一人サマースクールで知りあったのが、ショウジョウバエのホメオボックス遺伝子を研究していたマイケル・レビンです。体の背側を決める遺伝子のはたらきを調べ、動物の体づくりが共通の仕組みをもつことを示した人の一人です。彼も進化が気になっていて、ホヤの脊索の話をすると興味を持ってくれました。最初はピンチヒッターだった講師役ですが、次の年からレギュラーで呼ばれました。朝早くから夜遅くまで世界中の研究者と議論と実験を重ね、最先端の知識を得ながら、自らがひたすら実験することの重要性を学びました。それが研究なんだと。

サマースクールの講師を務めた米国ウッズホール海洋生物学研究所。左から3人目はウニの共同研究をしたエリック・デビッドソン博士。(左から2人目:本人)
脊索動物の形づくり
遺伝子を用いて発生と進化をつなげるという研究の方法論は見えてきたのですが、本格的な分子生物学をやるためのスタッフはいませんでしたし、予算の余裕がありません。最初は岡田先生の研究室で真壁和裕さん(現徳島大学教授)が実験させてもらい、乏しい研究費から少しずつ必要な機器を購入していきました。もし僕が死んで後払いの借金が残ったら大変だからと生命保険までかけたんです。学生さんにも満足な出張費を出せず、東北までホヤを捕りに行くのも青春18切符でした。それでも若い人たちが楽しそうに研究をしてくれたのが救いでしたね。
苦労しながらも、ホヤの脊索で発現する遺伝子を探し、その中にマウスの中胚葉と内胚葉の形成に関わるBrachyury という遺伝子があることを安尾仁良君(現フランスLe Centre national de la recherche scientifique 研究員)見つけ、これが脊索をつくるのに重要であることがわかった時は、具体的な方向が見えたと思いました。ナメクジウオの脊索でもこの遺伝子がはたらいていることを突き止めました。ところがです。なんと脊索を持たないウニやギボシムシにもこの遺伝子が存在したのです。なんだろう、この遺伝子は。そこで、さまざまな動物でのBrachyury のはたらきをよく調べてみたら、脊椎動物では脊索を含む中胚葉の形成と原腸の陥入に、脊索のないウニでは単に原腸陥入だけに関わっていることがわかりました。そしてホヤでは、脊索だけをつくってそれ以外の中胚葉の形成や原腸陥入には関与していないんです。ですから、ある動物からとったBrachyury 遺伝子が脊索形成能をもつかどうかを知るには、ホヤに入れて脊索だけをつくり出すことができるかをみればよいわけです。実際にウニのBrachyury 遺伝子は、ホヤに脊索をつくらせることができました。「脊索形成遺伝子」はいろいろな動物にあるのに、体全体をつくる発生の流れの中に脊索形成能が含まれるのは、脊索動物だけだったわけです。
脊索の研究が順調に進み、さらに脊索動物に共通の特徴であるエラや内柱(甲状腺の相同器官)の解析にも手を伸ばしました。こうして少しずつ、脊索動物進化の全体像が見えてきたと思っていたのですが、ある時、このまま続けて最後まで行ったら本当に進化がわかったことになるのだろうかと、ふと考え込んでしまったのです。脊索やエラは確かに重要な形質ですが、脊索のでき方やエラのでき方がわかることが本当に脊索動物の全体像を知ることになるのか。全体像を知るのが目的なら、まず遺伝子の全体像をつかみ、それから脊索動物の体制や進化の全体像を考えた方が早いのではないか。どうしてもゲノムの解析が必要だということになったのです。

研究室の新歓コンパにて。

試薬棚の前で。朝から晩まで実験三昧。
ホヤゲノムを読む
ゲノムプロジェクトは一大仕事ですから、ホヤゲノムを読みたいと一人で思っていてもできるわけはありません。日頃から、ホヤの研究をここまで進めたから後はゲノム研究をする他ないと発言していたら、ある時岡田先生がやってきて、「あんたゲノムが読みたいんやなあ」と吉川寛先生や小原雄治さん(現国立遺伝学研究所所長)といったゲノム研究者を紹介してくれました。メダカゲノムプロジェクト(2006年解読)やコケゲノムプロジェクト(2007年解読)の仲間に、ホヤを入れてもらえることができたのです。
ホヤではゲノム情報の蓄積が少なかったので、ゲノムDNAに取り組む前にまずはゲノムから転写される遺伝子配列の網羅的な収集を始めました。この計画を立てていたとき、まったく偶然なのですがアメリカでホヤゲノムの解析を始めるから加わらないかと、マイケル・レビンから電話がかかってきたのです。アメリカのゲノムセンター(JGI)のダン・ロークサーが力わざでゲノムDNAの断片的な配列をかき集め、その配列を僕のところで読んだ遺伝子情報と重ね合わせることで解読がスムーズに進みました。
2002年の4月、ゲノムの全体像が見えてきたところで、共同研究者が一堂に会してそれぞれ自分の興味のある遺伝子がゲノムのどの場所にあり、どんな構造をしているかを確認しあいました。9月に入り、この成果で論文発表をしようとダンと話がまとまりかけていたところに、遅れてやってきたマイケルがそれではダメだと議論をひっくり返したんです。「マウスやショウジョウバエなどのモデル生物とホヤでは、研究者人口の桁が違う。ホヤのゲノムはこうでしたという論文では価値がない。ホヤのゲノムを読んで初めて脊索動物の進化がわかるのだという内容でなければいけない」と言うのです。そしてさらに、多細胞生物のゲノムプロジェクトは他にもどんどん進んでいるから、一流の雑誌にホヤゲノムの成果を載せるには今年中が勝負と発破をかけられました。
僕はなるほどと納得しましたが、ダンはまとめ役としてのやる気を無くしてしまったようで、僕とマイケルの草稿があがっても彼の担当部分が全然出てこない。他の共同研究者から、どうなってるんだというクレームが出始め、マイケルは「俺はもう知らん」と言い出したんです。しょうがないので僕が仲裁に入って、みんなの利害を調整し、なんとかクリスマス前に論文が掲載されるその年最後の『Science』に間に合いました。同じ年にマウスゲノムやフグゲノムが報告されましたから、マイケルの言う通り脊椎動物ではない脊索動物のホヤゲノムの全容が同時に発表されたことに、大事な意味があったわけです。研究の成果を少しでも評価の高い雑誌に出し、注目されるかたちで発表するためのアメリカ流の執念や駆け引きに巻き込まれ、とても大変でしたが、めったにできないいい経験でした。もうやりたいとは思いませんけど。僕は体は丈夫なのですが、この時はさすがに終わった後に倒れてしまいました。

Scientist Library:
生命誌46号
『DNAのふえ方から見えた生きものの姿』
吉川寛

岡田節人先生(左)の文化勲章受章記念パーティーにて。

ゲノム解読後に開かれたジャンボリーに世界中の研究者が目当ての遺伝子を探しに集合。(前列左から2人目:本人)
ゲノムから発生と進化をさぐる
ゲノムを読んだところでまず僕たちのやることは何か。助教授の佐藤ゆたかさんを中心に、ホヤゲノムには、発生に関わるどんな遺伝子がどれだけあるかを洗いざらい調べました。シグナル分子はどれだけあるか、転写因子はどのタイプがどれだけあるかという内容で、10本の論文にまとめました。これは進化発生学の専門誌に投稿したのですが、この号は1冊まるごとぼくらの研究室の論文で埋まったんです。他の研究者もその論文をよく引用してくれ、そのおかげでどちらかというとマイナーなこの雑誌の知名度が上がったほどです。僕も、死ぬ時にお墓に何か1つ持っていくとしたら、研究室の皆で作りあげたホヤの報告づくしのこの雑誌ですというぐらい気に入っています。
こうして遺伝子のカタログができてみると、ヒトとホヤにあって他の動物にはないもの、ヒトにあってホヤにないもの、ホヤにあってヒトにないもののリストを作れます。脊索動物の中ではホヤが最も脊椎動物に近いのは確からしいのですが、リストを眺めると、動物のくせに植物の細胞壁を合成する酵素を持っているとか、前後軸に沿った体づくりのための遺伝子が壊れているとか、固着生活を送るホヤ特有の進化も見えてきました。こうなると、脊椎動物の系統からは離れているけれど、体のかたちはサカナにそっくりなナメクジウオのゲノムが気になります。ホヤゲノムを解析した国際共同研究のメンバーで、今ナメクジウオゲノムの解読を進めています。今度は僕は体を考えてあまり面倒見無いぞと言っているのですが。
ゲノムを読んだ結果できるもう1つの研究は、遺伝子の一つ一つが発生の時間の中でいつどの細胞ではたらきはじめるかを調べて、発生における遺伝子ネットワークの構造を明らかにすることです。ウニの発生ではエリックが一足先に見事な成果を出しています。ホヤでも、研究員の今井薫さんがやってくれました。初期発生でどの遺伝子がどういう順序ではたらくかという図をウニとホヤで比較すると、どちらもよく似たネットワークが見えてきます。ただ、ウニでは5時間かかることを、ホヤではたった5分で済ますという大きな違いがあるのです。この差を気にする人は少ないのですが、時間という本質的な問いが出てきたとわくわくしているんです。
最初にホヤゲノムを解析したいと言った時、小原さんから「ゲノムを読むということは、それが目的ではなく、ゲノム解読の結果をどう使うかが問題なんですよ」と散々言われました。そのときはピンと来なかったのですが、いまは小原さんの言った意味がよくわかります。ゲノムから発生と進化を探る試みは、これからが面白いのです。
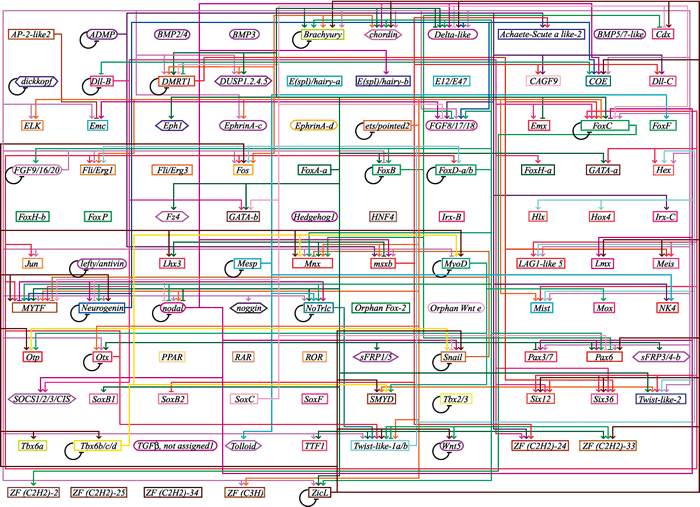
ホヤ胚の遺伝子ネットワークの概略。
カタユウレイボヤ統合型webデータベースより。

進化発生学の専門誌『Development, Genes and Evolution』2003年6月号。1冊が丸ごと研究室の論文で埋まった。

研究室のメンバーと大学の駅伝大会に参加。(前列左から3人目:本人)

雪国育ちなのでスキーは趣味。
200%の努力
僕自身はホヤを用い、発生と進化の2つの時間を見たいということにこだわってきました。それだけなのですが、ただ、仕事が何であろうと、持つ力の200%を出して取り組む姿勢は一貫しています。自分が立派な研究者だと思ったことは一度もありません。たまたま合格した理学部で生きものの面白さに気づき、良い師につくことができて良い仲間に恵まれ、気がついたら発生進化学の道を突き進み、ホヤ研究のパイオニアになっていた。今は、少しでも若い人がホヤ研究に参画しやすいように、縁の下の力持ちとして、ゲノムデータベースの公開や、バイオリソースとしてホヤの供給も続けています。
僕たちはとても幸せな世代だったと思います。発生学と進化学の分野で、ホメオボックス遺伝子の発見のようなわくわくすることが次々に明らかになってきた。研究者が仮に30年という時間を持つとして、その時代の研究の進歩にわずかでも貢献できたら、それは非常に喜ばしいことです。
大学の研究室は、学生を育てることが基本であるというのも、僕の一貫した方針です。研究テーマは相談して決めますが、それをどう展開させていくかは学生に一任しています。生物学の一番の楽しみは、誰もやっていないことを自分で見つけて展開し、皆に面白いと言わせることですから。この京都の研究室から、これまでに60人ほどが学位を取って巣立っていってくれました。これから研究者を目指す人には、きちんと自分で生きものを見て、色々なことを勉強して、本当に何をやりたいかを考えて欲しい。これが次の世代の人への僕のメッセージです。
(文責 今村朋子)



研究について語る。

ホヤの飼育室も兼ねた実験室。75歳くらいまで現役で研究したい。

ホヤとナメクジウオが脊索動物であることを明らかにした先人を記念してつくられたアレキサンダー・コワレフスキーメダル。賞に興味はないけどこれは嬉しい。

