



チョウと枯草菌
小学校4年生の時、先生からモンシロチョウの育て方を教わったのが生きものとのつきあいの始まりなんです。チョウ好きですが、卵や幼虫を捕ってきて育てるのが面白いので、珍しい種にこだわるのでなく、ありふれたチョウが成長していくのにじっくりとつきあうわけです。そうすると、このイモムシはキャベツが好きなのに、別のイモムシはミカン科の葉しか食べないということが分かってくる。多様性が面白いという見方が植えつけられました。今も続いています。
学生時代にDNA研究を中心にした分子生物学に触れ、そこに進みましたが、当時皆が研究していた大腸菌でなく、枯草菌を研究することになった。偶然です。注目されない研究対象を選んだことで、不愉快な思いをしたこともあります。しかし他人とは違う生物を研究をしたおかげで、DNAのふえ方という生きものの基本現象を大腸菌と比較することで、共通性と多様性がより深く理解できたと思います。チョウも枯草菌も偶然であった相手ですが、この二つを大事にしてきたことで、生きものの本質を考え続けることができたと思っています。

生後8ヶ月。
昆虫学者を目指す
小学校最後の年に終戦です。神戸の中心、六甲に住んでいたので、空襲を避けて山地の方に集団疎開していました。たった数ヶ月の疎開生活でしたが、栄養失調になりかけて、迎えに来た親がわからなかったぐらいです。家が焼けたのでとりあえず親戚の家に落ち着きましたが、しばらくは体を治すのに精一杯でした。それで1ヶ月くらい遅れて小学校に戻ったら、先生の最初の言葉が「来年中学受験やのに何しとった!」です。その先生が持っている教科書といえば、ついこの間まで使っていた本をスミ塗りしたもの。こっちは生きるだけで必死だったのに、先生は時代の変化なんかなかったように教壇に立っている。かなり反感を持ちましたね。
当時の中学には、まだ旧制時代の教養主義の伝統が残っていて、生徒が自主的に様々な活動をしていました。僕が所属したのは「六甲博物会」。中学のクラブと言っても、旧制の5年生もいたし高校生も参加していたから、新入生から見たら先輩は大人です。植物分類の名人とか、昆虫のことなら何でも知っているとか、かたつむりの渦巻きを数学的に解析するとか、すごい人がいる。そんな中でもまれながら、チョウの飼育に熱中していました。他のことではかなわないけど、チョウの飼育なら負けないぞという、子どもっぽい自己主張もあったかもしれません。そういうわけで、絶対に昆虫学者になるつもりで、京都大学の農学部を受けようと思っていました。ところが高校の修学旅行で東京に行き、泊まった旅館が本郷にあったので自由時間に赤門(東京大学正門)をくぐり三四郎池などを廻りました。すっかり東大の雰囲気が気に入ってしまい、先生からは今頃から勉強しても受からんぞと怒られましたが、なんとか理科2類に入りました。

国民学校1年生。母、姉、妹と神戸の自宅前で。

高校時代の昆虫採集(左端:本人)
生物学から生化学へ
大学で最初に出会ったのが松原謙一君(大阪大学名誉教授、DNAチップ研究所社長)です。彼はお父さんが植物学者で、ともに生物学を目指す仲間として意気投合しました。ある講義でボールドウィン(イギリスの生化学者)の「比較生化学入門」という本に出会い、生きものと物質をつなぐ「生化学」という分野があることを知り、二人ともたちまち魅了されました。化学は追試を受けるぐらい苦手だったのですが、生化学を学ぶには化学科しかないと考え進学しました。
化学の授業はやはり退屈で、図書館に行って生化学を独学し、生まれたばかりの全学連(全日本学生自治会総連合)に参加して過ごしていました。サンフランシスコ講和条約のあと、日本の社会が再び揺れ始めた頃です。ただし学生運動はまだおとなしい頃で、理学部の委員長となった僕の仕事は、全国行脚して各大学の組織と連絡体制を作ることでした。このとき知り合った研究仲間もたくさんいます。そういう時代ですから図書館で読む本も、オパーリン、ルイセンコ、エンゲルスとやはりソ連の影響が強かった。エンゲルスの「タンパク質は生命の本質なり」という言葉に我が意を得たりと思い、オパーリンの唱える物質から生命が誕生するという仮説に興奮しました。ルイセンコの獲得形質遺伝説もしばらくのあいだ完全に信じこんでいました。もちろんワトソン、クリックの業績はすでに知られていましたが、DNAは本質じゃないと思っているから全然頭に入ってこない。生化学を研究する前から、すっかり頭でっかちな生化学者の気分でしたね。将来自分がゲノムを研究することになるとは夢にも思いませんでした。

東京大学理学部化学科に進学。クラスメートに松原謙一君がいた。(後列左から2番目。後列中央:本人)
枯草菌と出会う
卒業研究は、日本の酵素化学の先駆けの一人赤堀四郎教授(大阪大学名誉教授、故人)の講座で行いました。赤堀先生はタンパク質の構造解析に取り組む傍ら、オパーリンのいう化学進化が液体(海水)と固体(粘土など)の界面境界で起こることを提唱し世界的に注目されるなど、あこがれの存在でした。卒業テーマはウマアルブミンのアミノ酸配列の決定で、ようやく生化学に触れることができ元気づきましたね。大学院はそのまま化学系に進みましたが、赤堀先生が当時できたばかりの応用微生物研究所(現分子細胞生物学研究所)の講座を兼任なさったので、面白そうだぞと思いそちらで研究を始めました。ここは理学・農学・薬学の各分野が集う学際的な研究所で、最新の機器もそろっていましたし、教授、助教授は、他の研究室と掛け持ちでほとんど不在。残った助手や大学院生は、「自由区」と呼んで、自分たちだけで研究を考えて楽しんでいました。
手の野村眞康さん(現カリフォルニア大学教授)や1年先輩の西村暹さん(現筑波大学監事)、大石道夫君(現かずさDNA研究所所長)、水野重樹君(東北大学名誉教授、故人)らと話し合い、タンパク質合成の仕組みを調べようと言うことになりました。モノーらが大腸菌を使って遺伝子発現のメカニズムを解き明かしつつあり、タンパク質合成にRNAが関わっていることはまだはっきりしていなかった時代です。モノーらは、大腸菌が異なる栄養源に置かれると、それに合った酵素を合成し生き延びるといういかにも生きものらしい仕組みを研究して、世界を驚かせたのです。
僕たちは、枯草菌が、増殖の段階に応じて作り出すタンパク質を大きく変えるという大腸菌にはない性質を持つことに着目しました。培養液中の枯草菌は、活発な増殖が一段落するとそれまでになかった様々な酵素を一挙に合成し始めるのです。細胞内には酵素の前駆タンパク質が蓄積されていて、増殖の状態が切り替わるとそれを酵素に変換するという仮説を立てました。そこで、増殖中の菌を放射性物質で標識し、どのタンパク質に標識が見られるかを調べればいいだろうと思ったのです。実は、タンパク質が1個1個のアミノ酸から生合成されるという、今だったら高校の教科書にあることも分かっていなかったのです。アイデア倒れでしたが、生命現象の素過程を探る方法論を試行錯誤できたのは、貴重な体験でしたね。

一年先輩の西村暹さん(左)と北海道旅行。
実験前につぶれたテーマ
大学院を出て、これからの研究課題を2つ考えました。1つは大腸菌の分子遺伝学。モノーを始めとした世界中の研究者が、DNAがどうやって表現型を生み出すのかを解明しようとしていました。もう一つは、その大腸菌に感染するウイルス(ファージ)について。アメリカのベンザーシーモア・ベンザー草創期の分子生物学を発展させた立役者の一人。多くの日本人研究者が影響を受けた。
参考
Scientist Library 堀田凱樹がファージの遺伝子地図を作り、遺伝子の実体がDNAの塩基単位にまで解析できる事を示しました。どちらも生命の本質を解く大事な研究です。最初は伝染病研究所(現医化学研究所)でウイルスの研究をやろうと思っていたのですが、医学系でポストを見つけるのが難しく、ポスドクに応募したら補欠の一番目でした。困っていたら、たまたま米国留学から一時帰国していた野村さんから、末岡登(現コロラド大学名誉教授)という若い研究者が独立してポスドクを募集中との話を聞きました。
末岡さんは木原均木原均植物遺伝学者。コムギの染色体倍加を研究し、ゲノム概念を確立した。
(1893~1986)の最後のお弟子さんで、渡米後はアカパンカビの遺伝学や密度勾配遠心法によるDNAの塩基組成分析を行うなど、古典的な遺伝学から最新の分子遺伝学まで精通した方でした。独立後のテーマとして遺伝暗号の普遍性の検証を考え、その材料として枯草菌に着目したのです。枯草菌は人工的に外来DNAを取り込ませることができるという、実験には好都合な性質を持っていました。形質転換と言います。遺伝暗号が普遍的でないなら、枯草菌に他の細菌のDNAを導入してもDNA配列は正しく読まれないでしょうし、普遍的であるならもとの細菌と同じアミノ酸組成のタンパク質が作られるだろう。遺伝子工学の技術がまだ確立していない時代に、斬新なアイデアです。急いで枯草菌の形質転換技術を学び、渡米しました。
1961年の7月、イリノイ大学の末岡研究室に着きました。まずは日本から持ってきた枯草菌の変異株を増やして、やる気満々、新しい実験の準備です。しばらくして、末岡さんはモスクワで開かれる分子遺伝学の国際学会に出かけました。その間もせっせと実験準備を進めていると、末岡さんからの手紙です。「ニーレンバーグマーシャル・ワーレン・ニーレンバーグアメリカの生化学者。遺伝暗号の解読とタンパク質合成の仕組みを解明した功 績により、1968年ノーベル賞医学生理学賞を受賞(ロバート・ホリー、ハル・ コラナと共同受賞)。が遺伝暗号の解読を発表した。僕らのテーマは意味がなくなったからやめる。次に何をやるか、考えといて」とありました。研究がめまぐるしく進展する時代にいるのは楽しいのですが、学会の発表一つでいきなりテーマがつぶれてしまったのにはちょっと参りました。

イリノイ大学に留学。大学のシンボル Alma Mater 像の前で夫人と。
ゲノムDNAの増え方
とにかく枯草菌を増やしながら新しいテーマを考えていました。ちょうどその頃発表された、生体内のタンパク質は端から連続的にアミノ酸をつないで合成されるという論文を読み、僕はゲノムDNAの複製も同じように順序よく合成されるのではないかと思いつきました。ゲノム上の決まった位置に複製の開始点があり、そこからDNA合成が始まって、最後に終結点にたどり着くと2つのゲノムになる。原核生物といえども丸ごとのゲノムDNAはタンパク質とは比べ物にならないくらい長い分子ですが、そのDNAが複製される時に、きちんと同じところから始まって同じところで終わると考えたのです。このような発想が出てきたのも、生きものを物質から見る時も必ず全体を考えて、そこに規則性を求めるという末岡さん流の研究スタイルの影響です。
このモデルが正しいとすると、どういう実験を組めば良いのか。増殖中の枯草菌の中で起きていることを調べようとした大学院時代の研究もヒントになりました。失敗した研究もいつか役立つもの。ゲノムを一つの連続した直線DNAと考え、端から順に複製されると予想すると、開始点付近のDNAは複製開始後すぐに2倍になり、逆に最も遠い部分は最後に2倍になるだろう。つまりDNA上にいくつかの目印をつけて、それらが2倍になる順番が決まっていることを示せればよいのです。
DNA上の目印とは、実際には遺伝子領域のことです。栄養要求性栄養要求性多くの野生型の微生物は、糖分および無機物の窒素源を含む培地で生育可能である。これに対し、特定の栄養素をさらに培地に加えないと生育できなくなった変異体を栄養要求性変異体と呼ぶ。このような変異体の多くは、特定の栄養素を合成する酵素が働かなくなったものである。の枯草菌の変異体をいくつか使って、どの遺伝子領域が2倍になったかを測定する方法を考えました。例えば培地にアデニンを加えないと生きていけないアデニン要求性の変異体に、野生株のDNAを導入すれば正常なアデニン合成酵素遺伝子を持ち、アデニンの入っていない培地でも生息可能になります。1回の形質転換操作でどれだけの変異体を野生型に戻せるかは、加えた野生型のDNA量に比例します。枯草菌は胞子を作るので、増殖が確実に止まっている状態のDNAを基準にすることができ、この実験のモデル生物としては実に好都合です。
激しく増殖している枯草菌の集団では、1つの細胞が分裂して2つに、さらにその2つが分裂して4つにと、指数関数的に菌数が増加しています。この集団をゲノム複製の進行状況からみると、全ての細胞で複製開始点付近の遺伝子は複製が既に終わっており、複製終点付近の遺伝子は未複製の状態です。つまり、開始点付近と終結点付近の遺伝子頻度は集団として常に2対1となり、その中間に存在する遺伝子は、開始点からの距離に応じて2から1の間の値をとります。この値を、形質転換効率として求めたのです。自然に増殖している細菌集団を用いてDNAの複製順序を推定できるという、画期的なアイデアでした。
指数関数的に増殖している野生株の枯草菌からDNAを回収し、12種類の栄養要求性変異株を野生型に戻すための形質転換効率を測定しました。幸運なことに、この12種類の中には、常に比率がほぼ2であるものと、つねにほぼ1となるものがあったのです。前者が複製開始点の近傍にある目印、後者は終結点の近傍にある目印になるということです。残りの目印も、ゲノム上のどの位置にあるかがわかりました。ゲノムDNAは全体がきちんとした順序で複製されていることをついに証明できたのです。留学から2年後のことでほっとするやら嬉しいやら。
この成果をコールドスプリングハーバー研究所の会合で発表しようとしたら、メセルソンマシュー・メセルソンアメリカの分子遺伝学者。スタールと共に、密度勾配遠心法によりDNAの半保存的複製を証明した。のグループが大腸菌で同じことを発表するというのです。しかし彼らはたった2個ほどの目印を調べただけで、こちらのデータの方が優れているという自負がありました。ところがモデル生物である大腸菌を使っているというだけの理由で彼らが先に発表し、こちらは脇役といったプログラムでした。末岡さん共々頭に来て、座長に発表順序を変えさせましたよ。

末岡登さんと枯草菌ゲノムの研究を始めた頃。イリノイの夏の一コマ
ゲノム上に目印を見つけて、ゲノムDNAの複製が進む順番を調べた。この図では、左が複製開始点で右が終結点を示す。adeは開始点のすぐ近傍にある目印であり、複製開始直後にすぐ2倍になる。
複製開始点取り競争
NASAがカリフォルニア大学に作る分子生物学の研究所に誘われました。なぜNASAが枯草菌かと思いますね。宇宙開発を進めていく中で、地球上の生物をむやみに宇宙施設や他の惑星に持ち込んではいけないという意識が出てきた。枯草菌の胞子は熱にも紫外線にも宇宙線にも強いので、これを滅菌できる条件を開発すれば生物汚染の心配はないだろうということで呼ばれたのです。一応これに沿った研究もしましたが、ほとんどはDNA複製の仕事を続けました。
一つは複製と細胞周期の関係です。細胞がDNAを合成する速度は、複製開始の頻度と複製伸長の速度によってきまります。このうち伸長速度は、栄養状態つまり細胞の増殖速度とは無関係に一定であることが分かりました。ですから、DNA複製の周期と細胞増殖の周期を調和するには、複製開始の頻度を調節しなければなりません。つまり栄養条件が良いと開始頻度を多くし、栄養条件が悪いと開始をとめることもあるのです。一方、普通の栄養状態のときに複製酵素の阻害剤を加えるなどして複製の伸長速度を遅らせると、相対的に開始頻度が増えたことになるので、次々に複製開始点が増幅したやまたのおろちのようなゲノムを作ることが出来ます。この図は複製開始と細胞周期の密接な関係を示すものとして、よく引用されました。
この現象を応用すれば、複製開始点のDNA領域を単離できるのではないかと考えました。やまたのおろち状態になったゲノムは、正常なゲノムに比べて何倍もの複製開始点を含む領域を持っていることになります。このような菌体を集めれば複製開始領域が濃縮されたDNAを抽出できるはずです。実際に通常と比べると複製開始点が20倍以上濃縮されていました。しかしまだ遺伝子組換え技術が確かでなかった頃なので、その程度の濃縮率では膨大なゲノムDNAの一部の領域を単離できませんでした。
1969年、金沢大学に新設されたガン研究所に呼ばれて帰国した後も複製開始点探しを続けましたが、組換え技術でDNAの一部をクローニングできるようになった時には、大腸菌のグループに先を越されてしまった。大腸菌のゲノムDNAを取り出して短い断片にし、それを複製開始点を持たないDNAにつなげて大腸菌に戻してやるという方法を繰り返すことで、200塩基対ぐらいの領域がDNA複製の開始に十分であることが示されたのです。もちろん枯草菌でも同じ方法を試みていたのですが、どうしてもうまくいかなかったのです。

複製のやまたのおろち。(上)枯草菌ゲノムが複製され始めた直後の状況。通常はこのままDNAが倍加する。(下)実験的に、DNA複製の速度が細胞分裂の速度を下回る状況を作ると、複製が終結する前に開始点からの複製が次々起こってしまう。この模式図を「複製のやまたのおろち」と呼んだ。

金沢大学がん研究所の仲間と、自宅の庭でバーベキューパーティー。この庭にはチョウの食草園も作った。
複製開始の共通性と多様性
競争に負けてがっかりしましたが、ここで止めたら本当の負けになる。大腸菌と同じ方法で複製開始点を取り出せないのには、必ず理由があるはずで、そこには重要な生物学的意味があるかもしれない。大量の胞子を一斉に発芽させることで、最初に複製される領域を標識して分離することには成功していました。こうなったらその場所のDNA配列を全て決定して、大腸菌の配列と比較してみることだ。ゲノムプロジェクトの考え方ですよね。今なら誰でも思いつくことだし、すぐにできることですが、DNA配列の解読が自動化されていないので根気のいる仕事でした。助手の小笠原直毅君(現奈良先端科学技術大学院大学教授)が頑張って、およそ1万塩基対の領域を漏らさず決定しました。当時バクテリアでは一番長い、ゲノムの連続した配列情報です。少しずつ時代を先取りし、技術のないところで苦労しました。
苦労した甲斐がありました。大腸菌と枯草菌は同じバクテリアですが、系統的には十数億年ほど前に分岐したきわめて縁遠い生物です。それが、複製開始点のDNA配列も、その配列に結合するdnaAと呼ばれるタンパク質も非常によく保存されているという予想外の事実を発見しました。もちろん違っているところもあります。複製開始点とdnaA遺伝子が、大腸菌ではそれぞれゲノム上の異なる領域に存在しているのに、枯草菌ではそれらが近い位置にある。両方とも、複製開始点に複数のdnaA結合配列があるのですが、枯草菌ではその配列が広い領域に散在して、大腸菌ではコンパクトにまとまっている。これで、大腸菌ならでたらめにゲノムを切っても高い確率で複製開始点を無傷で取り出せるのに、枯草菌ではできなかった理由がわかりました。
複製開始点とdnaAタンパク質という、DNAが増える時に必要な役者は共通しているけれど、それをどのように使うかは違っている。つまり共通性と多様性という生物の基本が、このような基礎的な現象ではっきりしたのは面白いことだと思います。実は大腸菌では、複製開始点は細胞周期の制御にそれほど重要ではありません。大腸菌は条件さえよければひたすら増殖する性質を持っていますが、複製開始の制御を甘くすることでそのような生き方が可能になっているのです。一方枯草菌は、条件が悪ければ胞子を作って一休みし、DNA複製の開始が細胞増殖を強くコントロールしています。多くのバクテリアの開始領域を調べると、その特徴は大腸菌型、シュードモナス菌型、枯草菌型の3つに分類できることが分かり、これらが祖先細菌から進化・多様化してきた道筋も見えてきました。複製開始点とdnaAタンパク質の組み合わせからバクテリアの進化モデルを提唱した総説は、大腸菌の研究者からも高く評価されました。
かつてモノーは、「大腸菌で真であることはゾウにとっても真である」と、分子生物学が普遍的な生命現象を研究する強力な手段であることを訴えました。しかし枯草菌をよりよく知るだけでも、共通性だけではなく多様性がこれだけ見えてくるのです。

枯草菌研究の片腕、小笠原直毅君(現奈良先端科学技術大学院大学教授)。大阪大学医学部遺伝学教室にて。
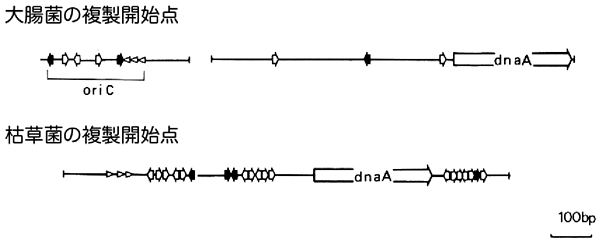
大腸菌と枯草菌の複製開始点の比較。短い黒矢印はdnaAタンパク質が結合する9塩基配列(TTATCCACA)を、短い白矢印はそれと類似した配列の存在を示す。またdnaAと書かれた矢印は、dnaA遺伝子を示す。大腸菌の複製開始点(oriC)は短くまとまっており、dnaA遺伝子とも離れた場所にある。枯草菌の複製開始点は、多くのdnaA結合配列とdnaA遺伝子を含む長い領域となっている。
枯草菌ゲノムプロジェクト
枯草菌の研究グループは、数が少ないこともあって世界中の仲間がよくまとまっています。競争相手と言うよりも、研究仲間の意識が強いのです。大腸菌グループに一致団結して対抗しなければならないという面も確かにありました。ところが1990年にフランスで開催された枯草菌の国際会議の席で、欧米のグループだけで枯草菌ゲノムプロジェクトを構想していると突然発表されたのです。アメリカのグループがまとまった研究予算を獲得できる見通しが立ち、ヨーロッパのグループがそれをあてにして始めたものでした。寝耳に水の僕は、ゲノムプロジェクトのような研究基盤を作る作業は、皆が参加しその成果を共有できる国際共同研究とするべきだと強く主張しました。
この提案が受け入れられ、早速日本の研究グループをとりまとめてなんとか予算を獲得したら、ヨーロッパの方はEUの支援を受けるのに難航し、アメリカのグループはプロジェクトそのものから手を引いてしまいました。これは研究者側の問題ではなく、アメリカ政府は研究予算の使い道を常に国益に沿うか否かで審査します。アメリカ一国の予算でゲノムプロジェクトを行うのは意味があるが、国際共同研究にアメリカ市民の税金を投入するわけにはいかないという判断だったようです。こうなると日本は責任重大です。僕がブリュッセルのEU本部に乗り込んで、日本の研究グループは枯草菌ゲノムプロジェクトを必ず遂行する意志があることを明言しました。なんとかEUの研究グループも軌道に乗り、1997年に全塩基配列を決定し終えました。これが僕の枯草菌研究の一区切りで、ゲノム情報をもとにした枯草菌の新しい生物学は、若い研究者に引き継いでもらおうときめたのです。

ヨーロッパでの枯草菌ゲノムプロジェクトの予算獲得を後押しするため、単身EUの本部ブリュッセルに乗り込んだ。

日本のゲノム研究を応援してくれた仲間達。左から、榊佳之(現ゲノム科学総合研究センター長)、松原謙一、藤山秋佐夫(現国立情報学研究所教授)、吉田光昭(現万有製薬株式会社つくば研究所長、本人)
複製開始点から見えた真核生物らしさ
金沢から大阪大学に移った時、ずっと気になっていた真核生物の複製開始点の研究を始めようと思いました。原核生物は染色体当たりに複製開始点が1つだけ存在するので、染色体そのものが複製単位(レプリコン)となります。しかし酵母菌などの真核生物では、1つの染色体に複数の開始点が存在することが知られていましたが、なぜそうなっているかはわからない。酵母菌の染色体は真核生物の複製モデルとして世界中の研究者が注目しており、新参者の僕がみんなと同じ事を考えていては勝てるはずはない。独自の視点は何かと原点に戻れば、僕達が枯草菌の複製現象をゲノム全体で捉えたように、やはり真核生物の染色体全体で複製現象を理解すべきだということです。
これまでの研究は、染色体から複製開始点を取り出してその性質を調べるというものばかりで、その結果、酵母染色体は複数の複製開始点を持っているがそれらの働きに違いはないとされていました。しかし染色体に存在する全ての複製開始点の位置を調べて、複製の時にそれぞれがどういうはたらきをしているかを調べた者はいない。染色体の全体像を見れば、きっと新しいことが見つかるに違いない。そこで入学したばかりの大学院生の白髭克彦君(現東京工業大学助教授)を説得し、酵母染色体の研究に着手しました。
幸いなことに、酵母ゲノムプロジェクトを始めたアメリカの研究者から6番染色体をカバーするゲノムDNAの断片をひと揃いもらうことができました。まず断片をつなぎ合わせて染色体の全体像を把握し、個々の複製開始点の分布を調べました。6番染色体は平均的な細菌ゲノムの10分の1程度の大きさしかないのに、開始点を10個も持っています。DNAを複製している時にそれぞれの開始点が活性化する順番を調べると、染色体の中心(セントロメア)に近いところから始まって、遅れて周辺部の開始点が活性化していました。複数の開始点を持っていても、結果的に染色体全体としては原核生物のように順番に複製が進んでいたのです。
また、複数の開始点の存在はDNA複製に必須ではなく、大半の開始点をつぶしても酵母は生育できました。一つの開始点から複製が始まれば、後は順番に進んでいくのだから当然と言えば当然です。ところがさらによく調べると、面白いことに染色体に散在する開始点は複製の開始を制御するのではなく、DNA複製時の異常を感知するとその位置で複製を停止させるはたらきを持つことが分かったのです。最初に活性化する開始点は、細胞増殖のシグナルを受けて複製を開始します。他の開始点は、何事もなければ遅れて活性化して複製を始めますが、DNAに傷が入るとそのシグナルを感知して、修復されるまで複製をストップさせるのです。できるだけ正確に染色体を複製しようとする真核生物の工夫が、1つの染色体に複数の複製開始点を持つことだったのです。染色体全体を見るアイデアでDNA複製の真核生物らしさを知ることができ、とても面白い研究ができたと思っています。
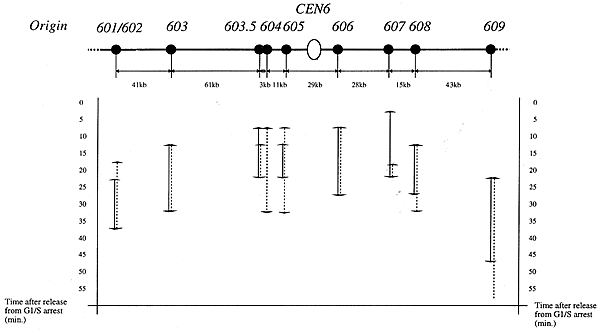
(上)出芽酵母第6染色体の構造。白丸はセントロメア、黒丸は複製開始点を示す。(下)細胞周期がスタートした時を0分とし、それから何分後の間にどの複製開始点でDNA複製が始まったかを、上図の複製開始点に対応する縦線で示した。複製は染色体の中心付近の開始点から始まり、両端の複製開始点は最も遅く始まる。
社会の中の科学者
これまでを振り返ってみると、社会が激しく動き、学問が大きく変わる時代を経験した気がします。小学校を卒業する時に終戦を体験し、学生時代は安保問題、在米中はベトナム戦争で揺れる社会を目の当たりにしました。そのため、世間で当然と思われていることを疑ったり、民主主義は素晴らしいけれどもいかにもろい制度かということを意識したり。アメリカを去る直前、カリフォルニア州知事が保守的な人物に代わり、リベラルなカリフォルニア大学の学長を罷免した時は学内が激しく混乱しました。抗議活動をする学生を州兵が狙撃した光景は、今でも忘れられません。
職業として科学者を選んだわけですが、研究さえしていれば良いのだと思ったことはありません。市民としての科学者のありかたを常に考えてきました。例えば自分たちが研究を続けられる状況をどう作ればいいのか。誰かが研究者の声を代弁するのではなく、研究者一人ひとりが考え、必要があればまとまって行動することも大事です。若い研究者の就職難は、半世紀前から問題となっていました。大学院の時に生化学若手の会を結成したのはそういう背景があったのです。僕は2代目の会長を務め、ポスドク問題に取り組みました。
一方、科学者の社会的責任を考え、自分の正義感をどう貫けばいいのか悩んだ時もあります。金沢にいたときはスモン訴訟スモン訴訟胃腸薬として投与されたキノホルム剤による薬害被害者の救済を訴えた裁判。国及び製薬企業の責任が追及された。の弁護団に加わり、スモン病がキノホルムによる薬害であるとの原告側の主張を支持する活動をしました。松枯れ対策としての広範囲の農薬散布が、実は全く無関係な虫を殺す効果しかないことを地道な調査で裏付け、行政に訴えかけて見直してもらったこともあります。科学者という専門職に就く市民として、その場その場でできることをしてきたつもりです。

元昆虫少年として、小学生と夏休み合宿を楽しむ。(生駒山:コスモスセミナーにて)
昆虫少年の分子生物学
日本に帰国して不満だったことの一つは、DNAの研究が物質と生命現象を結びつける分野だとなかなか認知されていないことでした。これは研究者どうしの交流が不十分であることにも原因があると思い、先輩や同僚と協力して日本分子生物学会を創設しました。
実は僕らの年代の分子生物学者には昆虫少年だった人が多いのです。大澤省三さん(前JT生命誌研究館顧問)もその一人でした。分子生物学会がなんとか軌道に乗りはじめると、DNAのことばかり議論するのも面白くないという愚痴が出てきます。そこで大澤さんらと分子生物学会「虫の会」を作り、学会がある度に元昆虫少年たちが集まるミニ集会を開きました。噂を聞きつけた岡田節人さん(同前館長)が、「学会には興味ないけど虫の会には入れてくれ」と現れ、蝶の話で盛り上がりました。そのつながりで、昆虫少年としての分子生物学をこの研究館でスタートすることができたのです。チョウはどうやって決まった植物の葉っぱに卵を産み、自分の子どもが食べ物に困らないようにしているんだろうという小学生の頃からの疑問に、若い人たちと取り組むことができてわくわくしています。
JT生命誌研究館の顧問として「昆虫と植物の共進化ラボ」を主宰。アゲハチョウ科の食草認識のメカニズムを探る

分子生物学の先輩末岡登さん(右)と大澤省三さん(中央)。

岡田節人先生と虫談義。





