

両親の背中を見て育つ
3歳の頃には、両親が営む千葉市内の街の本屋で店番をしていました。レジを打っていると、荒っぽく小銭を投げ出すお客もいれば、大人に対するのと同じように丁寧に接してくれるお客もいる。世の中には実に色々な人がいるということを、自然と受け容れていきましたね。この経験が僕の土台にあるのかもしれません。今思えば、植物について子どもや学生の頃から知りたかったことが、色々な人との出会いをきっかけに研究へとつながってきた気もします。
本が入った大量の段ボール箱を運んだり、伝票を作ったりと、常に働く両親の姿を見て育ちました。僕のワーカホリックな性質は両親に似たのかもしれません。父は独立心が強い人で、故郷の山梨県を出て一から店を開きました。配達帰りに立ち寄った喫茶店で、「人に使われる仕事は面白くないから、自分でやるのがいいんだぞ」と語っていたのを覚えています。母は幼いころ東京の家を空襲で焼け出されたり、疎開先で弟妹を亡くしたりと壮絶な経験をしたそうです。そんな苦労にも関わらず悲観的な面を見せたことはなく、どんな事も笑い飛ばしてしまう本当にすごい人です。今でも両親を世界で一番尊敬しています。
母は忙しい仕事の合間を縫って茶道と華道を習い、人に教えるまでになるほどで、自分ができなかったからと、僕には色々な経験をさせてくれました。切手やコイン、仮面ライダーカードの収集、エンジン模型飛行機作りや国体の鼓笛隊のトランペット奏者もやりましたし、ペットとして犬やジュウシマツ、タナゴ、さまざまな昆虫を飼育したり、習い事は絵画やそろばん、書道、剣道もやりました。中でも「あしのこ学校」というサマーキャンプは印象的でした。小学生から社会人までが一つのグループになり、キャンプをしながら議論して意見をまとめるというものです。年齢も職業も異なる人が対等に話し合えることが、子ども心に新鮮でしたね。色々な意見があっても、真摯に議論していくと、アウフヘーベン
アウフヘーベン対立する概念を、より高次のレベルで包摂すること。することがあるのだと知りました。母が経験させてくれたことはどれも、今も色々な場面で役立っています。

両親と旅行中の1枚。父も母も働き者で、根っからポジティブな性格だった。

色々なものをコレクションし、怪獣のソフトビニール集めも好きだった。研究者になって、奇妙な形の植物や突然変異体を目にしたときのわくわく感は、この頃ガメラを見て感じたのとあまり変わらない気がする。

休日には、家族で近所の田んぼにオタマジャクシを採りに行ったり、潮干狩りに行った。

毎年数回は、家族旅行や書店の親睦旅行に出かけた。家族旅行では、妹がぐずって列車に乗り遅れたり、台風で船が欠航したりと想定外のことが次々と起きたが、両親が機転を利かせて対応する姿を、子ども心にかっこいいと感じた。
食虫植物との出会い
母曰く、2歳の頃から植物が好きだったそうで、小学校の頃にユリの鱗茎鱗茎食用にする百合根。ユリの地下茎を葉が鱗状にとりまいたもの。に心ときめいたのを今でも覚えています。ただ、それを次元が変わるほど高めてくれたのは、小学校から高校にかけての同級生、鵜澤(うざわ)武俊くん(現 大阪教育大学教授)との出会いです。小学5年の時、彼が手作りの温室で育てていたハエトリソウやウツボカズラなどの食虫植物に、一瞬にして魅了されてしまったのです。さっそく僕も父に手伝ってもらって、ヒヨコ電球で暖める温室を作り、食虫植物を育て始めました。以来、互いの家をよく行き来するように。何を話すでもなく、子供部屋で互いが持ち寄った園芸雑誌「ガーデンライフ」や食虫植物の本を黙々と読みふけっていましたね。
鵜澤くんが入っていた食虫植物研究会にも入り、大人に混じってフィールドワークを楽しみました。食虫植物の研究者である小宮定志教授と助手の柴田千晶さんが中心になって運営する研究会で、愛知県の豊橋市にある湿地や、今は開発でほとんど無くなりましたが、千葉県の南部にあった食虫植物の群落を見に行けたのは素晴らしかった。さらには、当時まだ珍しかった、ドロセラ・アデラエというモウセンゴケの仲間を大きく育てて展示会に出品し、第1回目の「食虫植物研究会賞」に選ばれました。しかし僕は、「植物が勝手に育っただけであり、自分は受賞に値しない」という趣旨の手紙を送って、受賞式を辞退。環境さえ合えば植物は自ずと育つのだという考えは今も変わりませんが、当時の僕はそこに「自分はいったい何をしているんだろう」と疑問を感じたのです。この時期から、食虫植物の栽培への興味が薄れていきました。

小学校2年の夏休みの自由研究で作った初めての植物標本。

小学校の頃、ランドセルに入れて持ち歩いていた食虫植物の本。右の本は、熱を出して寝込んでいた日の朝に届き、読み終えた昼頃にはすっかり元気になっていた。左の本の著者の近藤勝彦先生には、大人になってモウセンゴケの分子系統解析で共同研究のお願いに伺った際に、サインをしていただいた。
植物分類学を目指す
当時、県立千葉高校は学園紛争の影響が残っており、受験ばかりを視野に入れて学問をしない授業はタブーだという空気がありました。それでも皆はちゃんと受験勉強していたわけですが、それを素直に間に受けていた僕は、高校を卒業してそのまま浪人生活へ。予備校に通うのも楽しくて、「嬉しそうにしすぎだ」と父に嗜められたほどです。1年で偏差値が60上がるという経験をしました。
後から考えて浪人して良かったなと思うのは、この時期に将来の研究分野を決めることになったからです。高校時代は、生物研究同好会を作ってさまざまな本を読んだり実験をしたりして、柴岡孝雄先生に刺激されて食虫植物の電気信号の研究に興味をもったり、コンラート・ローレンツさんに影響されて動物行動学をやってみたいと思っていました。浪人していたある日、同好会の顧問だった岩瀬徹先生から、県立博物館の準備室に寄贈された植物標本の整理を手伝わないかと誘われたのです。この時、近所の千葉港に面白い帰化植物が色々あると教えてもらい、さっそく自転車で港に行き、片っ端から植物を採集、出版されたばかりの『日本の野生植物』という図鑑で種名を調べ始めたら、これが滅茶苦茶面白かった。特に、それまで一緒に見えていたイネ科の花を解剖すると、種ごとに顕著な違いがあることに感動しました。そして、今思えばあまりにも漠然とですが、「植物分類学」をやりたいと思うようになったのです。僕の思いを知った父が、本の配達先で紹介していただいた千葉大学の西田誠先生(元 千葉大名誉教授)に進路を相談してくれ、東京大学に、岩槻邦男先生という気鋭の分類学者がくるから東大を受けなさいと言われたのです。

高校の生物研究同好会の仲間と。文化祭のポスターを横にするか縦にするかなど、ささいなことを徹夜で議論したりしたのは良い思い出。それぞれ異なった仕事についたが、今でも毎年集まっている。(本人:左から2番目)

浪人時代に調査した千葉港の帰化植物リスト。これが最初の論文となった。
植物三昧がはじまった学生生活
無事に東京大学理科二類に入学し、生物学研究会というサークルに入りました。ここで知り合った、シダを得意とする先輩の鈴木武さん(現 兵庫県立人と自然の博物館研究員)、同級の牧雅之くん(現 東北大学教授)と一緒に紀伊半島から屋久島まであちこち採集旅行に行き、すっかりシダに魅了されました。同じサークルには、林学の宮下直さん(現 東京大学名誉教授)、鳥類の仲岡雅裕くん(現 北海道大学教授)や甲虫の濡木理くん(現 東京大学教授)、水生動物の坂本竜哉くん(現 岡山大学教授)など、今でも生物学で活躍している生きもの好きが勢揃いしていました。春から秋にかけて数回、仲間たちとテント生活をしながら、三岩岳や天狗原山にいる生物を全て調べる「総合調査」を行い、植物だけでも数百に上る生物種をリストにして、手作りのガリ版の冊子にしました。今でも貴重な資料ですよ。3年になると、無事に第一志望だった理学部の植物学教室に進級。午前は講義、午後は夜遅くまで実験、朝から晩まで分子生物学、生化学、生理学、遺伝学など、植物に関する全ての分野を学べて、もう天国でしたね。植物分類の先生は大場秀章さん(現 東京大学名誉教授)や邑田仁さん(現 東京大学名誉教授)、矢原徹一さんなどの達人ばかり。実習で日光、屋久島、大雪山などの植物を存分に採集し、学生時代に一万点近く植物標本を作りました。自分で採集して標本にすると忘れないもので、一つひとつを今も覚えています。また、シダ談話会という研究者の集まりで、形態学者の今市涼子さん(現 日本女子大学理事長)や化石学者の西田治文さん(現 中央大学名誉教授)たちと知り合い、いまだに多くのことを学ばせていただいています。
大学院生の時に、研究室の加藤雅啓先生に連れて行ってもらったインドネシア調査も忘れません。初日に「この調査の目的は何ですか」と先生に尋ねると、「馬鹿もん、行ってから考えるに決まっとるやろ」と一言。人知を超えた熱帯雨林の自然を前に、その言葉の正しさがわかりました。人々の姿もカルチャーショックで、子供たちは日本なら考えられないような、激流や断崖絶壁で平然と遊んでいるし、大人たちは一週間後に船で迎えを頼んだのに二日も遅れてきました。それまでの自分の常識や行動規範とは異なった、しかしそれでもきちんと成り立っている世界があることを知りました。相手の背景にある思想や文化を理解して付き合うことを学ぶと共に、自分の枠組み・世界そのものが広がったように感じました。



フィールドで植物を探すのは大得意で、ムカシベニシダ(上:大学3年)とオニマメヅタ(下:修士2年)いう超珍品のシダを見つけてご満悦。

2016年、300万点の植物標本を所蔵する北京の中国科学院植物標本庫で調査をしていたところ、偶然開けた棚に見覚えのある標本が。大学4年の時に自分が採集した標本だとわかり、奇跡的な再会を記念して撮影。

博士課程1年のインドネシア調査で、地元ガイドの方と。肩から下げているのは巨大な着生シダ。
植物分子系統解析の第一世代
1980年代、植物の分類には形態的な特徴が用いられていましたが、異なる系統で似たような形が進化する「収斂進化」を見抜くのが難しく、その分け方は文献によってずいぶん違っていました。そんな時、長谷川政美さんの『DNAからみた人類の起原と進化』という本に出会い、分子系統学を知りました。これまで形態を比べても結論が出なかった、オランウータン、ゴリラ、チンパンジー、そしてヒトの類縁関係が、分子系統解析によって、統計的な確率を含めて明らかになる研究史が、生き生きと書かれていたのです。さらにこの頃、大澤省三さんや堀寛さん(現 名古屋大学名誉教授)が、生物の普遍的にもつDNA配列(5SリボソームRNA)を解析し、全生物界を包含する分子系統樹を作り上げました。塩基配列やアミノ酸配列なら、生物の類縁関係を統計的に推定することができる。ぜひ植物でやってみたいと思いましたね。でも当時は、植物からDNAを取り出すことすら至難の業。大学院試験の面接では、形態学のゼミでお世話になっていた原襄(のぼる)先生(元 東京大学名誉教授)に「そんなことが簡単にできると思っているんですか」と散々に言われました。でも生意気な若者だったので、さらにやる気が出てきたんです。
当時の植物学教室には、卒業研究は大学院で進学予定の研究室とは、別の研究室で行うという決まりがありました。ここで、サルモネラ菌の遺伝子解析を行っていた飯野徹雄先生(元 東京大学名誉教授)の研究室に行けたことが、実に幸運でしたね。直接指導をしてくれたのは助手の沓掛和弘さん(現 岡山大学名誉教授)で、最初に「研究者はビジネスマンですから、朝8時に来て夕方5時まで実験をしてください。5時以降は論文を読んだり書いたりする、知的活動をしてください」と言われました。沓掛さんに実験の方法を手取り足取り教わり、実験ノートの書き方や論文の読み書きの仕方まで含め、たった半年で実験と研究の流儀を身につけることができたのです。
晴れて大学院で岩槻先生の研究室に進みました。研究室がある小石川植物園の建物は古く、電気泳動槽にコオロギが泳いでいたこともありましたね。DNAを使って研究する人は僕以外にいませんでしたから、部屋の隙間を塞ぐことから始め、一から実験室を整備しました。岩槻先生は、大学院のテーマを相談に行ったら「やりたい事をやったらええで」とおっしゃって、後はセミナーで時々コメントをくれるくらいでした。でも必要な試薬や機器を買っていただいたことはもとより、助手の村上哲明さん(現 兵庫県立人と自然の博物館館長)と、アメリカの植物分子系統のラボを巡れるよう旅費を工面してくれたり、葉緑体DNAの第一人者である杉浦昌弘先生のもとに実験を習いに行く段取りをしてくれました。大学院生の教育の真髄を、身を以て学ばせてもらいましたね。おかげで植物分子系統解析の第一世代として、小石川植物園の研究室には、全国から共同研究者が集まるようになったのです。

小石川植物園の地下に整備したDNA実験室。整備前の写真が無いのが残念。
分子植物地理
学部学生の頃から知りたかったシダ植物の系統をすぐに調べたかったのですが、DNAがうまく取れません。ワラビやゼンマイのぬるぬるした食感からわかる通り、シダ植物は多糖類が豊富に含まれているため、DNAを取り出すのが難しいのです。そこでシダのDNA抽出と並行して、たくさん材料が手に入り、比較的DNAが取りやすかったカエデを使って修士の研究をはじめました。
19世紀、ハーバード大学の植物学者エイサ・グレイは、黒船が日本から持ち帰った植物が、北米のそれと酷似していることに驚いたそうです。太平洋を挟んだ日本と北米で植物相がなぜ似ているのかについては、二つの仮説がありました。一つは、かつて地球が温暖だった時代、北極に巨大な森があり、その森が寒冷化に伴って南下したため、北米にも日本にも同じ植物があるのだという説。もう一つは、氷期と間氷期を繰り返す間に何度も陸橋ができて、風や鳥によって種子が拡散し、大陸間を行き来したという説。僕はカエデ属の植物でこの仮説を検証しました。東アジアと北米に隔離分布している近縁なカエデのペアが分かれた年代を推定していくと、ペアごとに分岐年代が違っていました。つまりカエデの種子が何回も海を行き来したということであり、後者の仮説が支持されたのです。この成果を日本植物分類学会で発表したら、植物分類学者である清水建美先生(元 金沢大学名誉教授)が、「分類学会で始めての、分子生物地理の発表ですね」と褒めてくれたことを覚えています。
その後、世界各地に点在するモウセンゴケ属の分布や、横山潤くん(現 山形大学教授)や鈴木三男さん(現 東北大学名誉教授)と植物でもっとも顕著な隔離分布をするドクウツギ属の研究も行いました。これらと他のグループの研究からも、被子植物の分布はどれも大陸移動ではなく、長距離分散によって引き起こされた可能性が高いことがわかっています。


北米西海岸に生えるカエデの仲間Acer circinatum(上)。日本のコハウチワカエデ(下)と形態がよく似ている
想定外の系統推定結果
本筋のシダについては、100種以上のシダを調べてホウライシダの素性が良いということを突き止めました。ホウライシダの葉緑体のDNAを採って遺伝子地図を作り、葉緑体ゲノムの全制限酵素断片を大腸菌にクローニングしました。それまでシダの葉緑体DNAのクローンバンクはなかったので、世界から反響がありましたね。博士1年の頃に、有名なシダ学者のデビット・コナンさんから国際電話がかかってきて、「プロフェッサー(教授)の長谷部はいるか?」と尋ねるので驚きました。シダのDNAのクローンを分けてほしいという連絡だったのですが、「プロフェッサーかあ」と、ちょっといい気分でしたね。
博士過程では、自分で採集したシダ植物の、主要な属の系統解析を行いました。得られた結果は従来の仮説とは大きく異なり、目レベル以上の違いがあると考えられてきた、デンジソウとサンショウモが同じグループになってしまいました。岩槻先生も加藤先生も「長谷部くん、大丈夫かあ?」と半信半疑。しかし同じタイミングで、オハイオ州立大学のガー・ロスウェルさんが、デンジソウとサンショウモの共通祖先にあたる化石を発見したのです。アメリカのシダ研究の大御所ウォーレン・ワグナー先生が「クラッシクな論文になるかもしれない」とおっしゃって、米国科学アカデミー紀要に推薦してくれたのは幸運でした。後に留学した際は、この論文が話題になってあちこちの大学にセミナーに呼んでもらい、アメリカ人コミュニティーの一員になれたことが、帰国後にとても役に立ったのです。
シダと並行して解析した裸子植物の系統も、意外なものでした。この頃、植物園に来ていた植田邦彦さんや伊藤元己さん(現 東京大学名誉教授)たちと、裸子植物の形態進化について夜な夜な議論したのですが拉致があかず、DNAで系統をはっきりさせることになりました。ところが、現生の裸子植物が単系統になってしまったのです。つまり、針葉樹やイチョウやグネツム類など、葉の形も生殖器官の形もすごく異なる植物が、同じ祖先から派生したという結果となり、誰も信じてくれなかった。実は10年後にも同じことが起こったんです。ポスドクだった西山智明くん(現 富山大学特任准教授)が中心になってコケ植物の分子系統解析を行なったところ、なんとまた単系統に。形態のかけ離れたセン類、タイ類、ツノゴケ類が共通祖先から派生したことになり、案の定、誰も信じてくれませんでした。結局、前者の裸子植物については台湾の盟友である趙淑妙さん(シューミャオ・チョオ:現 台湾中央研究院卓越研究員)らによって、後者のコケ植物についてはゲノムプロジェクトなどによって、どちらも発表後10年近く経ってから正しさが証明されました。

DNAが抽出しやすかったホウライシダ


デンジソウの仲間(上)とサンショウモの仲間(下)。シダ植物は、デンジソウ目、サンショウモ目、シダ目の3つに分かれると考えられてきたが、分子系統解析の結果、デンジソウ目とセンショウモ目は姉妹群でシダ目の中に含まれてしまうことがわかった。

学生時代に針葉樹類に近縁という予想外の結果になった、グネツム類のキソウテンガイと記念撮影(ナミビアのナミブ砂漠)。

研究室の助手として引率した、屋久島宮之浦岳の学生実習にて。左から本人、当時学部生の三宅崇さん(現岐阜大学教授)、加藤雅啓助教授(現東大名誉教授)。
分子系統解析からの新たな問い
このように分子系統解析の結果は、形を基準とした従来の分類を、予想外に覆すことになりました。僕は植物の形態が好きで、アーネスト・ギフォード先生とエイドリアンス・フォスター先生が著した『維管束植物の形態と進化』という教科書を翻訳して形態に精通していたこともあり、分子系統解析を行なった当時は自分自身が一番驚きました。解析は実験から統計処理まで自らの手でやっているので手違いはないはずなのですが、形態学者としての僕はとうてい信じることができなかった。一体、生物の形はどのように進化するのだろうという新たな疑問が湧いてきたのです。
ドイツのマックス・プランク研究所のジェフ・シェル先生が、東大の植物学教室にセミナーで来てくれたことが転機になりました。彼はキンギョソウという左右相称の花が、たった1つの遺伝子の突然変異によって、アサガオのような丸い放射相称の花になるという話をされました。遺伝子一つの変化で植物の形が劇的に変わると知って、目から鱗でしたね。さらにこの時、『Plant Cell』という雑誌のエッセイに驚くべきことが書かれていました。花をつくる「ABCモデル」の主要な遺伝子である「MADS-box遺伝子」が、花をもたないシダ植物の、リチャードミズワラビにもあるかもしれないという、未発表の結果が紹介されていたのです。形の進化の研究に転換するには環境を変えた方が良いと思っていたので、「花の咲かない植物で花の進化を探る」というテーマを瞬時に思つき、これで日本学術振興会の海外特別研究員に応募することを決めました。さっそく、エッセイで紹介されていたパデュー大学のジョー・アン・バンクスさんに、そちらで研究したいとファックスを送りました。次の日に届いた彼女からの返信はたった一行、「ノープロブレム」。こうして、植物の形の進化を探る研究に舵を切ることになったのです。

学生の頃のバイブルだったアーネスト・ギフォード
花の咲かない植物の花遺伝子
パデュー大学に着くと、さっそくジョディ(バンクスさんの愛称)が、リチャードミズワラビにMADS-box遺伝子があることを示すオートラジオグラムを見せてくれました。「ほら、ここにバントがあるでしょう」と言われて見ても、何も無い。こうやって見るのよと、フィルムを斜めにしてみせてくれても僕には何も見えず、本当なのかと不安になりましたね。自分で実験してみようにも、台湾の留学生チークワン・ウェンくん(現 中国科学院教授)に、「ねばねばしてDNAはほとんど取れないぞ」と言われる始末。でも学生の頃からの経験を活かし、DNAとRNAの抽出は半年で何とかクリアしました。しかし肝心のMADS-box遺伝子はその片鱗すら見つからず、1年半、さまざま実験を行いました。ネガティブな実験結果が出た寒い夜に限って、頑丈な鍵がレンチで切られて自転車が消えていたりと散々でしたが、ラボのポストドクは「ウエルカム トゥ ユナイテッド ステイツ!」と茶化してくれるし、僕もなるようになるだろうと楽観的な日々でした。
MADS-boxは、被子植物では花を作る遺伝子です。ある日、リチャードミズワラビにこの遺伝子があるとしたら、少なくとも、葉や生殖器官が分化する前の新芽の段階で発現しているはずだと思いつきました。でも新芽の原基は1本から数mgしかとれません。そこで、借りていた温室のスペースをこっそりと毎日少しずつ拡大し、温室一杯になるまで鉢を増やし、妻に手伝ってもらって全ての株から新芽を切除しました。1週間後、ついにパソコンの解析結果に、そらで覚えていたMADS-boxの配列が出てきたのです。すぐにジョディにメールすると、数秒で「コングラチュレーションズ!」と一言返信がきました。嬉しかったはずですが、もはやよく思い出せません。花の咲かないシダ植物に、花の遺伝子が見つかった瞬間です。

花の咲かない植物で初めてMADS-box花形成遺伝子の相同遺伝子が見つかったリチャードミズワラビ

ジョディと珍しく実験打ち合わせ。彼女は、朝、「みんな、なんか新しいこと見つかった?」とラボに来て、あとは幼稚園の娘の面倒を見ながら自分一人で実験をしていた。実験について議論することはほとんど無かったが、MADS-box遺伝子が取れた後、メイヤロヴィッツさんのところで実験するきっかけを作ってくれた。

実験の待ち時間が苦手だったので、最大7つの実験をうまく組み合わせ並行して行っていた。


妻との植物探索旅行。アメリカ滞在中は、実験の合間に妻と二人で北米、南米各地へ、いろいろな植物を見に行ったことが一生の宝になった。ガラパゴス諸島(上)の単純な植生を見て、進化論を考えるのにうってつけの場所だと納得できた。サボテンは日本で栽培していたが、アリゾナの自生サボテン(下)には度肝を抜かれた。
ヒメツリガネゴケをモデル植物にする
留学から植物園に戻り、MADS-box遺伝子の研究を続けました。リチャードミズワラビのMADS-box遺伝子がどこで発現しているのかを詳しく調べると、被子植物では機能分化しているA、B、C遺伝子に相当する遺伝子が、どれも同じ場所で発現していることがわかりました。シダ植物では花を作る遺伝子は機能分化しておらず、発現場所の変化が花の進化を引き起こしたのだろう。こう推測はできたのですが、現状では、発現している組織で本当に働いているのか、働いていたとしてもどんな働きをしているのかがわからない。論文を書きながらフラストレーションを感じました。遺伝子の機能を確かめるには形質転換をしなくてはならないのですが、リチャードミズワラビではどんなに頑張ってもうまくいきませんでした。
そんな時、偶然にも他の研究室から居候していた学生の藤原研二郎くんが使っていた、ヒメツリガネゴケに出会いました。容易に遺伝子ターゲティングができ、遺伝子と形の進化を知るのに最適な生物です。幸運にもこのコケに興味をもった大学院生の西山くん、榊原恵子さん(現 立教大学教授)、日渡祐二くん(現 宮城大学教授)たちと共に基礎生物学研究所(基生研)で一から実験系を確立しました。基生研の村田紀夫先生が、自身の主催したシンポジウムに、ヒメツリガネゴケ研究の開祖のデビット・コーブさんや、同僚のアンディ・カミングさんを呼んでくれ、彼らと深く議論し、国際コミュニティーの一員になれたことも研究を進める上でとても役立ちました。さらに、毛利秀雄所長が特例として雇用してくれた博士研究員の小藤累美子さん(現 金沢大学助教)が参入し、ヒメツリガネゴケのMADS-box遺伝子の機能解析を始めました。しかし遺伝子を壊しても何も起こらない。さまざまな試行錯誤の末、大学院生の越水静さん(現 遺伝学研究所助教)が、この遺伝子がコケの茎の長さを制御していることを突き止めるまでに、20年かかったのです。MADS-box遺伝子以外にも、シロイヌナズナやイネでは発生に欠かせないとされる重要な遺伝子をいくつも欠失させて機能を推定したのですが、どれも被子植物とは機能が全く違う。モデル植物と同じ機能の遺伝子は論文になりやすいのですが、違う機能だと、それが生物学的に面白くない限り、有名な雑誌には掲載してくれません。動植物の発生進化研究の全体像を理解して一般性のある問題点を把握することが必要になります。
この点で、基生研で研究できたことがたいへん役立ちました。基生研には動物の研究者と植物の研究者が同居しています。基生研に移った当時は、発生プロセスから進化を理解しようとする学問分野「進化発生生物学(エボ・デボ)」が、動物と植物で同時に芽生えつつありました。どちらも多細胞体制をもつ生物同士、その発生の違いを知ることは重要だと思い、エボ・デボを一緒に考える日本初の国際シンポジウムを開催。植物発生分野から塚谷裕一くん(現 東京大学教授)、動物発生分野から阿形清和さんを共同オーガナイザーとして誘いました。先導的な研究を行っていた海外研究者に加え、日本の植物と動物の発生学者の多くが集まってくれました。多くの動物研究者と知り合い、彼らとの議論を通して植物の特徴を再認識するとともに、動植物をあわせて、発生進化学の一般性のある問題点を整理することができました。

ジョディは、家族とともにサバティカルで1年間基生研に滞在し、さまざまな共同研究を展開することができた。西表島で、ゲノムを解読したイヌカタヒバと記念撮影。


ヒメツリガネゴケ。大腸菌のシャーレで培養でき、酵母と同じ効率で相同組換がおこる。基生研で確立した実験プロトコールはインターネットで公開し、国際スタンダードとなった。また、隔年で開いた国際トレーニングコースには海外からたくさんの研究者や学生が自腹で参加した。

左はヒメツリガネゴケの野生型で、葉と葉がほぼ接しているので、毛細管現象で株元の色水が下の葉から上の葉へと運ばれ、先端部で受精がおこる。MADS-box遺伝子が無いと葉と葉の間が広くなり、水が運ばれず受精できない。被子植物以外ではじめてのMADS-box遺伝子の機能解明となった。ほとんどのコケが密に葉を付ける理由がわかって面白かった。

日本初のEvoDevoシンポジウムのポスター
国際ゲノムプロジェクトから見えた植物の姿
2000年になるとミレニアム・プロジェクトとして、文科省のゲノム特定領域研究が始まりました。ちょうどヒメツリガネゴケのゲノム解読の国際コンソーシアムを立ち上げた時で、西山くんと藤田知道くん(現 北海道大学教授)が中心となり、関原明さん(現 理化学研究所チームディレクター)、篠崎一雄さん(現 理化学研究所グループディレクター)たちに完全長cDNAライブラリー、藤山秋佐夫さん(現 遺伝学研究所特命教授)、小原雄治さん(現 遺伝学研究所特命教授)たちにその配列決定やEST解析をしてもらいました。
ゲノム解読ができると研究室の全員で、それぞれが興味をもつ遺伝子から始めて被子植物の発生に関わる500近い遺伝子の系統解析を行いました。すると驚いたことに、重要な発生遺伝子に相当する遺伝子の多くが、小葉植物のイヌカタヒバや、コケ植物のヒメツリガネゴケには無いのです。あったとしても、遺伝子数が大きく異なっていました。例えば、MADS-box遺伝子は被子植物では約40あるのに、イヌカタヒバでは3つ、ヒメツリガネゴケは6つしかない。これでは被子植物と同じ機能を担うことは難しいはずです。個々の発生遺伝子を見ていた時から、陸上植物は系統ごとに異なった発生プログラムを持っているのではないかと考えていましたが、ゲノムの情報もそれを支持していました。
動物の場合、プラナリアもヒトも、形は違っていても基本的なボディプランは同じですが、植物の場合、コケも小葉植物も被子植物も平らな葉をもっているのに、それをつくるしくみは違っているようです。植物は雌雄同体で自家和合性のことも多いですし、倍数体になって遺伝子が増えることも多く、突然変異が固定しやすいのかもしれません。その結果、いろいろな遺伝子ネットワークが独立に進化して、光合成に最適な平らな葉を進化させてきたのではないかと考えています。


ヒメツリガネゴケ国際ゲノムコンソーシアム。良く飲み、良く語り、良くメールし、国際共同研究の醍醐味を満喫できた。中央が本人、左から3人目が西山智明くん。

基生研の毛利所長が中心となって蝶類DNA研究会を設立し、会務や蝶の系統解析のお手伝いをした。2003年にJT生命誌研究館オサムシ研究グループと合併し、昆虫DNA研究会が設立され、現在に至っている。子どもの頃から昆虫採集や飼育が好きだったこともあり、多くの昆虫研究者と共同研究や交流を持てたことで研究の幅が広がった。写真は、2003年12月に開かれた昆虫DNA研究会とDNA考古学研究会の合同シンポジウム。

エリオット・メイヤロヴィッツさん(現 カリフォルニア工科大学教授)と。留学から帰る直前、彼の研究室で一ヶ月ほど実験させてもらい、その経験を活かして、帰国後、いろいろな植物に応用できるin situハイブリダイゼーション法を確立することができた。2025年に中国の学会で会った時、「おまえは一ヶ月うちのラボにいたから、いちおう同窓生だな」と言って、彼の研究室で活躍した伊藤寿朗さん(現 奈良先端科学技術大学院大学教授)と一緒に写真を撮ってくれた。
陸上植物の発生に共通性は無いのか
化石記録からは、陸上植物の共通祖先は、二叉に分岐する単純な茎のみからできた植物であり、コケ植物、小葉植物、シダ植物、種子植物でそれぞれ独立に複雑な茎と葉が進化したと推定されていました。僕たちが行った発生遺伝子の研究はまさにその仮説を支持していました。次の問題は、単細胞や糸状の体をしていた緑藻類から、どうやって陸上植物の共通祖先が進化してきたかです。裸子植物の単系統性に「オオ、ナイトメアー!」と言い続け、さんざん議論して仲良しになった古植物学者のピーター・クレインさん(
この答えは思わぬところから得られました。MADS-box遺伝子の研究に行き詰まった小藤さんが、造卵器と造精器の形成に重要な遺伝子の研究をはじめました。丁度、院生だった日渡くんが遺伝子トラップラインを作っていたので、造卵器と造精器に特異的に発現するラインを探し、トラップされている遺伝子を探しました。そこで見つかったのが、GRAS転写因子でした。研究は一筋縄にはいかず、結局、20年近くかかってしまいましたが、このグループの遺伝子を壊すとなんと、造卵器、造精器、葉脈ができなくなってしまったのです。
動物細胞と違って、植物細胞は細胞壁で覆われていて動けませんから、発生過程で細胞がどちらの方向に分裂して伸長するかで将来の形が決まります。単純な体制をもつヒメツリガネゴケは、発生過程の細胞動態をライブ観察できる利点があります。研究室を立ち上げて数年後、細胞の微小管をこよなく愛する村田隆さん(現 神奈川工科大学教授)に助教授として研究室に加わってもらい、細胞分裂の分子機構とその進化を研究室のテーマとして位置づけることにしました。
村田さんと毎日顔を合わせていると、自ずと細胞から形を見る習慣が身につきます。細胞生物学者の目で、造卵器、造精器、葉脈という異なった発生過程を持つ器官の共通性が見えてきたのです。どれも、組織の表面に平行な細胞分裂(並層分裂)が表皮の内側に細胞を作っており、できた内側の細胞が生殖細胞になったり水を通す細胞になっているのです。植物細胞は分裂面の面積が最小になるように分裂するというデフォルトルールがあるのですが、数理生物者の藤本仰一さん(現 広島大学教授)、鎌本直也くん(現 広島大学特任研究員)に細胞ごとのデフォルト分裂面を推定してもらったところ、GRAS転写因子は、このルールをやぶって無理やり並層分裂を制御する働きをしていることがわかりました。つまり、クレインさんの言う、植物の陸上化に必須だった器官や組織の進化は、GRAS転写因子の進化で説明できてしまうのです。さらに驚いたのは、GRAS転写因子は、土壌性バクテリアから陸上植物の祖先藻類に水平伝播したものだったことです。バクテリアの遺伝子の水平伝搬が、現在の陸上植物の体制を進化させた原因になっていたのです。
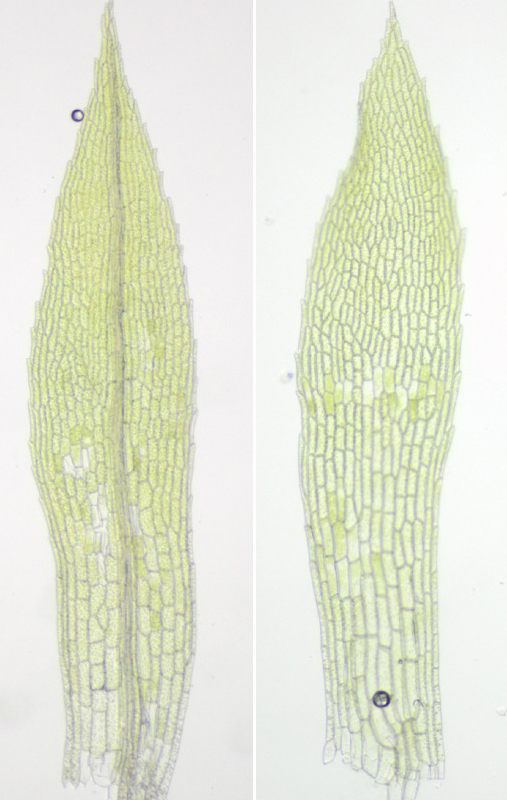
野生型(左)は葉の中央に葉脈ができるが、GRAS遺伝子を欠失すると葉脈ができなくなった(小藤累美子博士提供)。
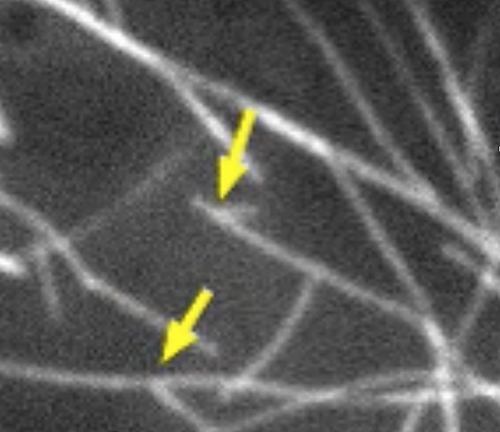
動物の細胞分裂では、中心体から微小管が伸びて紡錘体を作るが、植物の場合、紡錘体はできるのに中心体がないため、どうやって紡錘体ができるかは大きな謎だった。村田さんが蛍光タンパク質で微小管を可視化し、既存の微小管から新しい微小管が生じていることを発見し(黄色矢印)、植物では中心体が無くても紡錘体ができることがわかった(撮影:村田隆博士)。

壁谷幸子さん(元 基礎生物学研究所技術職員)が技術職員仲間の協力を得て、葉原基(上2枚)や造精器原基(下2枚)などの3次元細胞模型を作ってくれたおかげで、並層分裂(表面に平行な分裂)が見えてきた。赤い線が並層分裂面。葉原基の赤く色を付けて赤矢印で示した細胞が葉脈細胞、造精器原基の1N2bと書いた細胞が精子になる。銀色の丸いものは磁石。
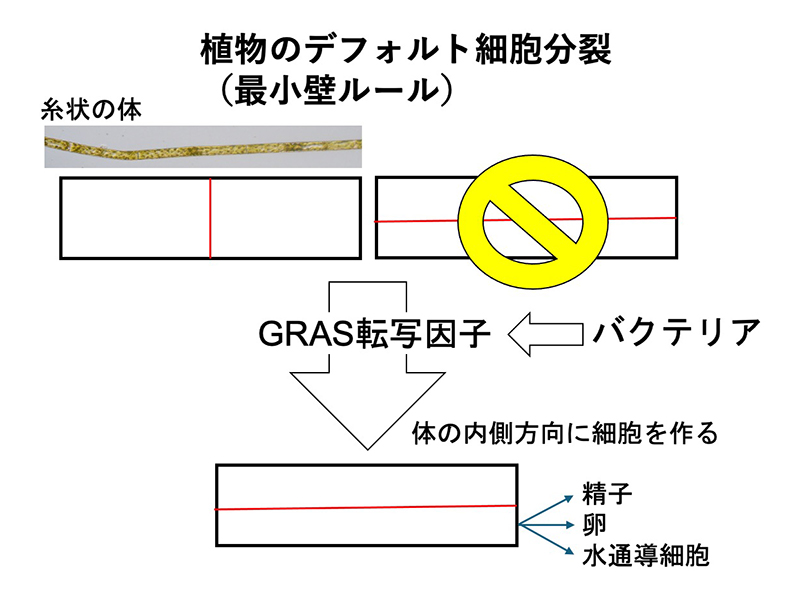
バクテリアから水平伝搬したGRAS転写因子が、植物のデフォルト細胞分裂を打ち破り、並層分裂を引き起こすことで、陸上生活に必要な造卵器、造精器、水通導組織が進化した可能性が高い。
植物進化の残された大問題
子どもの頃から、自分が負けるのも人が負けるところを見るのも嫌いで、今でもスポーツやゲームは好きになれません。競争にならないよう、他の人がやっていない研究ばかりしてきました。おかげで分子生物地理、分子系統学、発生進化、どれも第一世代として分野を切り開くことができたと思っています。これらに加え、植物の進化を考える上で欠くことができないけど、うまく答えの出ていなかった4つの問題について、従来とは異なる植物や実験技術を使うことで、競争なく研究を進めてきました。分化全能性、形態の大進化、短期記憶、そして電気信号です。
分化全能性の進化
植物は、挿し木でクローンが簡単に増やせることからわかるように、いちど分化した細胞が、容易に多能性幹細胞に変化します。このような植物の分化全能性は前世紀から知られていましたが、分化細胞が未分化な状態に変化した後、どの細胞が幹細胞になるのかを見極めるのが難しかったのです。分化全能性の綺麗な実験系ができないかと思っていた折に、思わぬところからチャンスを得ました。研究所で学生向けに実験生物のムービー集を作ることになり、ヒメツリガネゴケの再生過程を撮影することにしたのです。日頃から、ヒメツリガネゴケの葉を切って再生増殖を行なっていたのですが、改めてムービーを見て度肝を抜かれました。葉を切断すると、切り口に面した細胞が約1日で幹細胞に変化し、新たな植物体がにょきにょきと伸びだしてきたのです。これなら、葉が切断されてから幹細胞になるまでの遺伝子動態を追えるのではないか。ムービーを持ってERATOの面接に望んだらうまく採択してもらえました。5年間のプロジェクトの最後の年、ついに樋口洋平くん(現 東京大学准教授)と佐藤良勝くん(現 名古屋大学特任准教授)が、生体内で分化細胞を幹細胞に変えてしまうという想定外のパワーをもった、STEMIN転写因子を発見しました。研究を引き継いだ石川雅樹くん(現 基礎生物学研究所助教)たちが、STEMINはクロマチン構造を変化させることで、さまざまな遺伝子の転写を調節していることを見出したのです。
また偶然にもアメリカの友人に頼まれて、ヒメツリガネゴケのCSP1CSP低温ショックドメインタンパク質(Cold Shock Domain Protein)の略称。大腸菌が低温に順応する際に作られるタンパク質として研究されてきた。植物や動物にも共通した構造をもつタンパク質が見つかっており、低温だけでなくいろいろな環境変化によっても誘導されることがわかってきている。というタンパク質を調べていた佐藤くんたちが、これがヒメツリガネゴケで幹細胞を誘導することに気づきました。しばらく後に哺乳類の研究で、iPS細胞化に重要なLin28というタンパク質が発見され、なんとCSP1と相同な遺伝子だとわかったのです。院生の李琛くん(リー・チェン:現 湖北医薬学院教授)と助教の玉田洋介くん(現 宇都宮大学准教授)たちが解析を進めると、植物と動物の幹細胞化のしくみの違いが見えてきました。動物と植物では幹細胞をつくる際、クロマチン修飾や転写因子のネットワークを用いるという基本的なしくみは似ているけれど、働いている因子はCSP1以外は全く異なっていることがわかったのです。
残った疑問は、植物はどうして動物より幹細胞化しやすいのかという点です。これも偶然に、共同研究をしていた陈春丽さん(チェン・チュンリー:現 華中農業大学教授)の院生だった顧南さん(グ・ナン:現 四川大学卓越副研究員)が、ヒメツリガネゴケはDNAが損傷すると、分化細胞から幹細胞になることに気づきました。植物は紫外線や高温などから逃げ出すことはできません。そこでDNAが損傷すると、体の一部を幹細胞化して新しい体を作って伸びだし、より良い環境へ逃げ出すことが適応的だったのでしょう。これが、植物で容易に幹細胞化が起こるしくみが進化し、維持されている理由ではないかと考えています。
形態の大進化
2つ目の問題は、形態の大きな進化です。食虫植物の袋型の捕虫葉は、平らな葉からどうやって進化できたのか。捕虫葉は平らな葉よりも光合成効率が悪いのですが、獲物から栄養を取れる点で適応的です。しかし、捕虫葉に進化しかけの段階では、周りの植物より光合成効率が劣る一方、まだ獲物から栄養を捕ることもできないので、適応度が下がって進化できないはずです。小進化の積み重ねだけではうまく説明できない「大進化」はどうやって起こるのか。進化学に残された大問題の一つとして、倉谷滋さん(現 理化学研究所客員主管研究員)、深津武馬さん(現 産業技術総合研究所首席研究員)らと新学術領域を立ち上げました。運よく、食虫植物好きの福島健児くん(現 遺伝学研究所准教授)が大学院生として入学し、捕虫葉の原基では、表側基部で特徴的な並層分裂が起きていることを見つけました。基生研で数理生物学が得意な藤田浩徳くん(現 基礎生物学研究所助教)とともにシミュレートすると、原基での並層分裂による歪みが成長するにつれて大きくなり、袋状の葉ができることがわかりました。また、福島くんは食虫植物の消化酵素遺伝子は、普通の植物が菌類やバクテリアを殺すための耐病性遺伝子の相同遺伝子であることも見つけました。酵素の生化学的解析には朋友の鵜澤くんも加わり、共同研究として発表することができました。原基の特定の場所で細胞分裂方向を変えるだけで袋型の葉ができ、もともともっている祖先的な因子を消化酵素として使うことで、適応度をそれほど下げずに、大きな進化をしていくことができそうだと納得しました。
植物の記憶
3つ目の問題は植物の短期記憶です。ハエトリソウは、葉の上に生えた感覚毛に獲物が触れると葉を閉じ、獲物を捕まえます。しかし感覚毛に1回触られただけでは葉を閉じず、30秒以内に2回触られると葉を閉じます。つまり、1回目の刺激を30秒間だけ覚えていられるのです。隣のラボでマウスの短期記憶の研究をしている椎名伸之さん(現 基礎生物学研究所准教授)と立ち話をしていた時ふと、ハエトリソウは神経も脳も無いのに、どうやって覚えているんだろうと興味が湧きました。ちょうど、中学校のころに食虫植物の自由研究で相談に来たことがある須田啓くん(現 埼玉大学助教)が大学院に入学してきて、さらに運良く、北海道で開かれた内藤コンファレンスで、カルシウムシグナルを専門とする豊田正嗣くん(現 埼玉大学教授)に出会って意気投合。ハエトリソウにカルシウムセンサーを入れてみようということになりました。須田くんの2年半の奮闘の結果、ハエトリソウにカルシウムセンサー遺伝子を導入することに成功したのです。
ハエトリソウは感覚毛を1回刺激すると、まず葉全体にカルシウム波が伝播しますが、その1回だけでは葉を閉じる閾値を超えることはなく、2回目で閾値を超えることがわかりました。また、1回目の刺激によるカルシウムシグナルは時間と共に減衰するので、30秒以上たつと2回目の刺激でも閾値を超えないこともわかりました。動物の記憶形成と驚くほど似たシステムだったのです。
植物の活動電位発生伝播の分子機構と進化
小5のときに鵜澤くんの家で初めてハエトリソウを見た時以来、知りたいと思っていたけど最後まで残っていた疑問は、植物の電気信号、すなわち活動電位の発生・伝播のしくみとその進化でした。ハエトリソウの記憶の研究で印象的だったのは、カルシウム波の伝播速度が、活動電位の伝播速度とほぼ同じだったことです。真野弘明くん(現 基礎生物学研究所研究員)たちが並行して研究を進めていたオジギソウでも同じ結果が得られていました。実は、植物に動物の神経細胞のような活動電位があることは、チャールズ・ダーウィンがハエトリソウを、友人の電気生理学者に送ったことで見つかったのです。その後、植物の活動電位について多くの研究が行われてきましたが、遺伝子操作のできるモデル植物では、活動電位に加え色々な電気信号が混在するなどの理由で、活動電位に関わるイオンチャネル遺伝子は一つも特定されていませんでした。しかも、植物は動物で脱分極を引き起こすナトリウムチャネルを持っておらず、電気生理学的研究から、塩化物イオンの流出で脱分極すると推定されていて、新規性を秘めた現象だと感じていました。しかし、ハエトリソウは組織が複雑なため、活動電位の研究をするのは一苦労です。そこで注目したのが、食虫植物のモウセンゴケです。
モウセンゴケの葉には、粘性のある消化液を分泌する触毛がびっしり生えていて、獲物が触毛先端に触ると、触毛が葉の中央に向かって曲がり、獲物を包み込み消化してしまいます。50年ほど前に活動電位の研究がされており、触毛の先端部で活動電位が発生して柄を伝播、基部に伝わると、そこで運動を引き起こすことが知られていました。触毛の柄は2層の細胞があるだけなので、カルシウムイメージングと電位測定を同時にできるのではないかと考えました。ここでも苦労しましたが、瀬上紹嗣くん(現 基礎生物学研究所助教)、大井祥子さん(現 基礎生物学研究所技術職員)が学生や技術支援員たちとともに試行錯誤して、モウセンゴケの遺伝子改変と同時測定の実験系を確立できたのです。ハエトリソウとオジギソウのカルシウム波伝播のムービに希望を託して特別推進研究に申請し、無事採択してもらうことができました。モウセンゴケは大学院生の刘慧(リウ・フイ)さん、陈鹏(チェン・ペン)くん、2人を指導する张列弛くん(ザン・リエツ:現 基礎生物学研究所特任助教)と瀬上くんが、オジギソウは真野くんが解析を進めています。両種とも従来の仮説とは異なり、活動電位は細胞内へのカルシウムイオンの流入で引き起こされ、それを動物の神経細胞のシナプスで働いているグルタミン酸受容体と共通の遺伝子が担っているらしいことがわかってきたのです。定年までの1年半で、分子機構と進化の核心を明らかにできるのではないかと思っています。

ヒメツリガネゴケの葉を切って水に浸しておくと、切り口に面した細胞が1日ほどで幹細胞化し、2日ほどで伸び出す。切断後41時間後の写真(撮影:村田隆博士)。

ERATO「長谷部分化全能性進化プロジェクト」の発足時のメンバー。半分くらいの人数の基生研の研究室と同じ建物で連携しながら運営した。
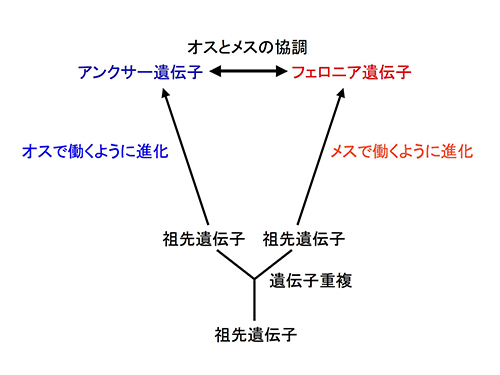
岡田典弘さんを代表とする「種形成の分子機構」特定領域研究で、宮崎さおりさん(現 静岡大学准教授)たちと遺伝子重複によってできた2つの遺伝子がシロイヌナズナの花粉(オス)と雌しべ(メス)で協調的に働くことで受精が可能になることを発見できた。この領域立ち上げのお手伝いをする過程で、岡田さんから面接のネクタイの色まで、いろいろなことを教えていただいたことが、自分が代表の新学術領域立ち上げに大いに役立った。


食虫植物サラセニア(ムラサキヘイシソウ)は壷型の葉で小動物を捕獲、消化、吸収する。たくさん葉を付けた自生株(上)と1枚の葉(下)。

北米の湿地に自生するハエトリソウ。
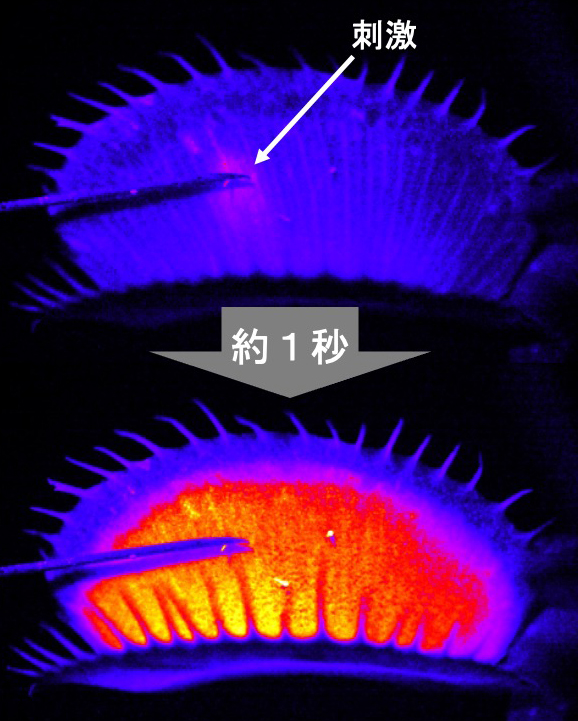
ハエトリソウの葉に刺激を与えると、刺激を与えた組織から葉全体にカルシウム波が広がることがわかった(須田啓撮影)。

ユーラシア温帯の湿地に広く分布するモウセンゴケ。コケではなく、ハエトリソウに近縁な被子植物。

獲物を捕らえたモウセンゴケの葉。葉に生えた触毛先端に獲物が触れると触毛が曲がり獲物は葉の中央に運ばれ消化されてしまう。

触毛の赤色色素が自家蛍光を発し共焦点顕微鏡での観察が困難だったが、野生集団から単離した赤色色素が減少した変異体を用いることがブレークスルーとなった。触毛先端を赤外レーザーで刺激すると、先端から基部にカルシウム波と活動電位が伝播し、触毛が曲がる。
陸上植物を体系的に理解したい
今までの研究を振り返ると、それぞれの研究を共にした人の顔と声が浮かびます。岩槻先生は「エデュケーションは教育やないで。人が育つのを妨げんことや」と言いました。まさしくその通りで、僕は学生と共同研究はしますが、これをやれと命じたことはないように思います。一緒に研究する中で何が必要かを話し合うだけです。それは子育ても同様で、子どもを見習って自分が変わったことは多くとも、子供に何かをするように言ったことはありませんね。植物のように、育ちたくなる環境をつくれば自ら育っていくものなのかなと思います。
学生が自由にやりたいことをやる研究室の方式を「放牧型」と呼ぶことがありますが、僕の場合は、自身の知りたい事が研究室運営の中心にあります。相手と自身の興味が重なり合った時、研究が躍進するという経験を何度もしてきました。僕はこれを「惑星直列型」と呼んでいます。自分の興味という必然と、人との出会いという偶然の両方から、思いがけない答えに辿り着いてきたのかなと思います。
僕自身は、実は今が準備万端、出発点なんです。分類学は、分類(Taxonomy)と体系化(Systematics)という二つの分野にわけられます。前者の分野では新種を記載することが主な作業になりますが、僕は後者の体系化をずっとしたかった。浪人の頃、分類学から植物の世界に入ろうと思ったのは、植物を総体として理解したかったからです。これまで、33万種強ある陸上植物について詳細な記載が行われていますが、情報源があちこちに分散していることと、情報量が多すぎることで、人間には全体像を理解することが困難です。一方、分子系統学で系統関係が明らかになり、種の個別的な記載を、系統樹という地図の上に配置することで関連性をもって理解できるようになりました。また、発生進化学とゲノム生物学の進展で、進化のしくみと素材がわかってきたので、個々の種の形質がどのように進化してきたかを推定することもできます。全体像を体系的に理解できれば、伝統的な形態学、古生物学での問題点を整理し、何が残された問題なのかを見つけることも可能になります。
体系化という仕事は今だからできることなんです。若い頃に何回か試みたのですが、経験と知識の不足で満足なものができなかった。今はフィールドに出ても、見えるものが若いころと全然違う。ずっと多くのものがそれぞれ連関して見えるんです。既に、目レベルで進化を体系的に考える本は作りました。これでうまくいった方法を、世界に1万5000ある陸上植物の全ての属に拡張した、植物図鑑を作ってみたい。従来の図鑑のように同定のための記載を羅列するのではなく、系統樹に基づいて、どこでどんな鍵形質が進化したのかを語っていく。「千一夜物語」のようにできないかなと思っています。これまで、だいたい150回、合計6年ほど出かけた海外出張の折に、世界中で約10万枚の植物写真を撮ってきました。また、自宅と山小屋で2000種ほどの植物を育て撮影してきました。だいたい主要なものは見たかなと思うのですが、細かい点ではまだまだ足りません。今は形態学全盛時代の論文のアーカイブスや、世界中の人が協力して作った植物写真のデータベースもありますから、それらで資料を補えば十分な図解ができるのではないかと考えています。毎日こつこつ続ければあと20年で完成できる予定で、その後5年はそれをアップデートしながら楽しみたい。これからが研究の本番でわくわくしています。



植物観察をしていたら小さな子どもが近寄ってきて(上)、同じように観察を始めた(下)。同じ興味を持った学生と教員が偶然出会い共同研究する、これが大学院教育の原点かなと感じた瞬間。

現生植物の目レベルの系統樹を出発点に、どの段階で進化の鍵となる形質が進化し、その後の進化に寄与したかを考察した教科書。植物のゲノム生物学、細胞生物学、発生学、形態学の知見に古生物学の知見を融合し、包括的に形態と進化を議論した点で、世界に類の無い本だと思う。出版後、何人かの気鋭の若手から、内容を十分理解した鋭いコメントや書評をもらった。他の人にも役立っているようだ。

ニューカレドニア固有の光合成をしない寄生性の針葉樹。どんな植物も、実物を自生地で見ると、文献からは気づかなかった形質に気づいたり、さまざまな疑問が湧いてきたりして、好奇心が爆発する。

研究所から車で40分くらいの場所に妻が見つけてくれた山林を購入し、野生動物の食害と戦いつつ、いろいろな植物を栽培し成長過程を撮影している。子どもたちが小さい頃は、良い遊び場にもなった。

