

工作に夢中になった少年時代
山形の農村部で育ちました。生家は祖父の代から医院を開業しており、幼い頃の私は診療所からもらってきた空き箱や絆創膏を材料に工作をして遊んでいました。ハサミで自分の指を切ってしまった時、誰にも言わず一生懸命に傷を隠していたと後年母から言われました。知られたらハサミを取りあげられると思ったのでしょう。一人で黙々と遊ぶのが好きでした。近所に幼稚園のような集まりがあり、そこに通うことになったのですが、大勢の子たちと一緒に遊戯などをするのは苦手だったんですね。最初の日のお昼に「いそがしいから、かえる」と言って家に戻ってしまいました。両親も無理に行けとは言わなかったので、結局それっきりです。人見知りで内向的な性格でしたね。
小学生の時に、親戚のお兄さんがボール紙で灯台の模型を作ってくれました。スイッチを入れて中の豆電球がピカッと光った瞬間、一気に心が惹かれました。本物の灯台みたいに光をぐるぐると回したいと、小さなモーターを使って自分であれこれ工夫しました。それ以来、車や船など様々な模型を作りましたが、条件は「動く」こと。飾るだけの模型には興味がありませんでした。

小学校の入学式の日に母と。のどかな地域で学習塾などなかったが、そろばん教室に通った。

診療所で診察をする父。私も医者になるのかなと小さい頃から思っていたが、中学生ごろの反抗期には数学者や物理学者に憧れていた。
留学で広がった自分の世界
田舎ですので、夜には家のまわりに明かりが少なく、見上げれば満点の星でした。中学生になり一眼レフカメラをもたせてもらうと、夜空にレンズを向けました。三脚に固定してシャッターを開け放すと、星の軌跡を同心円状にとらえることができるのです。診療所にあるレントゲン写真用の暗室で自分で現像しました。時々、同心円ではない軌跡が写り込みました。人工衛星ですね。どんな写真が出来上がるかと待っている時間も楽しみでしたね。星の明るさによって写真に映る光跡の線の太さが違うことに気がつき、幅を測って何等星か導き出す方法がないか解析を試みたこともあります。習うのではなく、いつも自分で工夫しながら遊んでいました。
学校の授業で好きだったのは数学です。基本の公理や定理さえ覚えれば、必要な式をそこから導け、答えが一意に定まるというのが私に合っていました。理科も好きでしたが、生物は苦手でした。おしべやめしべの数、多年草か一年草か、種まきと開花の時期など暗記することが多いですよね。論理的に答えにたどりつくのが好きだったので、自分なりにまとめの表を作り、何とか法則性はないものかと探しましたが挫折しました。それに山形は寒いから植物の時期が教科書に書いてある標準より遅かったり早かったりするのです。日常感覚と違うことを暗記しなければいけないことも不満でした。
高校3年生の時、学校の先生が交換留学を勧めてくださいました。米国で1年間ホームステイをしながら現地の高校に通うというものです。両親に相談してみたところ父に反対されてしまいました。父は、医学部卒業後に大学で研究をしたいと思っていたようですが、戦時中だったため軍医として中国に送られ、終戦後もシベリアで3年ほど抑留されていました。帰国した時には、祖父の医院を継ぐことになり、結局研究はできませんでした。戦争によって人生を回り道したと感じていた悔しさから、私には早く自立することを望んでいたようです。諦めかけましたが、医師をしていた16歳年上のいとこが、良い経験になると父を説得してくれたおかげで試験を受けることができ、3年生の夏にカルフォルニアへと旅立ちました。最初は言葉に苦労しましたが、気がつくと英語で日記をつけ、英語の夢をみるようになっていました。若い時は吸収力がありますからね。留学前の私はアメリカ人に対して西部劇に出てくるような武骨な人たちの集まりというイメージをもっていましたが、現地でそれが覆りました。非常に率直でハッキリと意見を言うけれども、繊細な気遣いもする。自分も彼らみたいなコミュニケーションの力を身につけたいと思うようになったのです。社交的になることで自分の世界が大きく広がることを初めて実感しました。この経験がなければ私はとっつきにくい嫌な大人になっていたんじゃないかな(笑)。人生の転機となった1年でした。
留学先で卒業証書をもらい、帰国して高校の一つ下の学年に合流しました。受験生ですが、土産に買って帰ったビートルズの「ホワイトアルバム」やクリームのレコードをかけて夏休みの間ずっと部屋でゴロゴロしていました。アメリカぼけからなかなか抜け出せなかったのです。母は心配していましたね。事実、漢字が書けなくなっていて、古文や漢文には苦労しました。でも、英語の成績は上がりましたし、化学はアメリカの教育がとてもよかったので応用して問題が解けるようになっていました。このように、留学が有利にはたらいた科目もあって、なんとか東北大学の医学部に進学できました。

高校での留学先、米国カルファルニア州ポトーラ市近くの山中で。

留学を終え日本へと発つ日。左:高校生の交換留学を主催する国際教育交流団体AFSの会長(当時)
力強い心臓の姿に惹かれて
医学部の授業は、全科目必修です。暗記しなければいけない事柄も膨大ですが、避けて通るわけにはいきません。自分の興味に応じて授業を選択できないことに、息苦しさを感じていました。そんな中で一番興味をもったのが生理学です。私たちのからだはどのように動き、はたらいているのか、生命現象を機能の面から捉える視点が工作少年だった私に合っていました。からだの構造の各部の名称を覚える解剖学は、暗記嫌いの私には難関と思っていましたが、比較解剖学を研究されていた助教授の佐藤達夫先生(現・東京有明医療大学学長)の講義のおかげで楽しく学べました。私たちのからだの神経、筋肉、血管、臓器のそれぞれの並びや位置関係には共通祖先から経てきた進化の時間が反映されており、構造に進化的な意味があることを教えてくださったのです。研究室を覗くとサメなどの解剖をされていました。
病院での実習が始まり、胸部外科の手術に立ち合う機会を得ました。この時、初めて拍動する心臓を直に見てその力強い動きに圧倒されました。休むことなく弛緩と収縮をくり返し、命を支える心臓に興味をもった私は、卒業後の進路として循環器内科を考えました。6年生になり内科の瀧島任先生(東北大学名誉教授、故人)にお話を伺いに行ったところ、「うちは内科医を10年計画で育てる。最初の4年は基礎医学の大学院生と同じように研究をしてもらう」と言われました。医学には、治療や診断に直接関わる臨床医学の他に基礎研究である基礎医学があります。当時の私は基礎医学に魅力を感じながらも、研究の世界が想像できず、自分が基礎医学に向いているという自信もなかったのでその思いを胸にしまっていました。瀧島先生の言葉を「内科医になるにしても、4年間は基礎医学の研究をしてもいい」と捉え、循環器に関わる基礎医学の研究室を調べ始めました。インターネットなどありませんから、図書室に行って論文を読み、自力でいろいろ探しました。全国どこでも訪ねてみようという心づもりでいたところ、同級生が骨格筋の世界的な研究者が東北大学に着任されたと教えてくれました。骨格筋は心筋と同じ横紋筋です。身近にそんなに素晴らしい先生がいらっしゃるなんて幸運だと、新任の遠藤實先生(現・東京大学名誉教授)の研究室の門を叩きました。この出会いをきっかけに、筋収縮そしてカルシウム・シグナル研究へと入っていくことになったのです。

大学ではバスケ部に所属していた。上下関係もそれほど厳しくなく、週3日ほど練習に励んだ。
学問の楽しさを知る
筋肉の収縮の制御を担っているのは細胞内のCa2+(カルシウムイオン)濃度で、例えば心臓では拍動ごとにCa2+濃度上昇が起きています。今では常識となっている、この「筋収縮のカルシウム説」を世界で初めて発表されたのが江橋節郎先生(東京大学名誉教授、故人)です。遠藤先生は江橋先生の門下であり、筋肉研究の大家アンドリュー・ハックスリーアンドリュー・ハックスリー【Andrew F. Huxley】(1917−2012)イギリス出身の生理学者。1954年に横紋筋の収縮のしくみを説明する「滑り説」を提唱した。イカの巨大軸索を使い活動電位の計測、解析を行い、1963年「神経細胞の末梢および中枢部における興奮と抑制に関するイオン機構の発見」によりノーベル生理学・医学賞を受賞している。先生の研究室への留学経験もお持ちでした。研究室に入るとさっそく遠藤先生から、ハックスリーの1957年の論文を手渡されました。前半は膨大な研究のレビュー、後半は数理モデルによって筋収縮のしくみを考察している非常に難解な内容でしたが、読み進むうちに次第に夢中になっていき、「世の中にはこんなことを考えつく天才がいるのか!」と衝撃を受けました。振り返ると、大学までの勉強は次のステップに進むためだけに必要な道で、興味がある科目だけに集中してとことん向き合う時間はありませんでした。一つのテーマに集中する自由な時間を得て、こんな幸せなことがあるだろうかと思いましたね。遠藤先生の下で、学問の楽しさを初めて知ったのです。大学院に進むまでは臨床医になる未来も考えていましたが、自らの問いに徹底的に向き合う研究の魅力に触れ、抜け出せなくなりました。
大学院に入って1年程たち、博士論文の研究テーマを決める時は、骨格筋か平滑筋かという選択に悩みました。運動神経の支配下にあってからだを動かす骨格筋は、素早く力を出すことに長けています。当時の医学生物学研究の中でも骨格筋研究は花形の一つでした。一方、消化管や血管、気管など内臓を動かす平滑筋は、骨格筋より未開拓な研究分野で、やりがいがありそうだが不安要素も多いと感じました。先生に伺ってもどちらがよいかをはっきりとはおっしゃいません。迷った末に、平滑筋への挑戦を決めその収縮に必要なCa2+濃度を探る研究を、実験系の確立から一人で行いました。3年かけて成果がまとまり、当時生理学研究で最も権威ある雑誌『The Journal of Physiology』に掲載できた時は、とてもうれしかったですね。遠藤先生の下で行った研究ですので、論文には先生のお名前を入れるのが普通です。ところがこの時、遠藤先生は「君が全部一人でやった成果だから単著にしなさい」とおっしゃったのです。自分が指導する立場になり、先生のふところの深さを改めて実感します。

アンドリュー・ハックスリー博士(左)と江橋節郎先生(右)。大学院で偉大な先生方の仕事を知り自分が研究社会で生きていけるのか物凄く不安になったが、やがて天才と同じことを目指さず、自分の道を進もうと考えられるようになった。
ロンドンでの2年間
次のテーマを考え始めた時、当時の平滑筋の生理学的研究では解析できることに限りがあり、さらなる技術開発をしないと前に進めない険しい道が続くと感じました。そこで再び骨格筋に眼を向けると、最も進んでいる研究が二つありました。一つは筋肉がATPの化学エネルギーをいかにして収縮という力学エネルギーに変換するのか、筋肉の機械的性質を探る研究です。もう一つは、神経からの刺激によって筋肉の細胞膜に起こる電位変化を細胞内のCa2+濃度変化に変換し、収縮につなぐ興奮収縮連関の研究です。考えた末、前者に取り組むことにしました。ハックスリーの弟子ボブ・シモンズ(Bob Simmons)博士がロンドン大学でこの研究を行っていたので、遠藤先生が手紙を書いてくださいました。すると、「私たちの研究室では実験装置を電子回路から手作りしている。故障したら自分で修理しなければならないが、医学部出身者にそれができるか?」という返事が届きました。医学部にそんな科目はありませんから、断りの手紙だとしか思えません。がっかりしていたら、遠藤先生が「それなら工学部に行って勉強してきなさい」とおっしゃったのです。前向きにとらえていいのだと驚きましたね。すぐに先生が知り合いの工学部電子工学科の助教授に頼んでくださり、4ヶ月ほど研究室でハンダ付けの方法に始まる電子回路の基礎を学びました。この時身につけた電子回路の知識は、留学期間のみならず、私の研究人生の中で大いに役立つことになりました。
ロンドンでの1年目は、先輩のポスドクと一緒に実験を進めました。収縮している筋肉に、小さな長さ変化を1万分の2秒というわずかな時間で加えて、筋肉が出す力の変化を見る実験です。先輩が標本づくりを担当し、私が実験装置のボタンを押すなどしていましたが、実験結果は芳しくありませんでした。標本をきれいに作り直したらどうかと言っても、なかなか聞き入れてもらえないのです。大学院の時は一人で実験しており、納得がいくまで試行錯誤できたので、今まで味わったことのないもどかしさでした。このままではダメだと、自分の意見を論理的に伝えるよう努力したことが、英語のとてもいい訓練になりましたね。データの解析にコンピューターを活用するなど、当時の日本の研究室にない手法を身につけることもでき、大きな糧になった2年間の留学でした。ただ心残りは、この間の成果を論文にまとめられなかったことです。妻と息子と3人で一緒に帰国する日を決めた後、どうしても最後まであきらめることができず、もう少し実験がしたいと一人でロンドンに数日残って実験を続けるということまでしたのですけれどね。3歳の息子を一人で連れて帰った妻には散々文句を言われました。

英国留学の夏休みに家族とアイルランド旅行に出かけた。

ロンドン大学を離れる直前、仕事場での一枚。

2度目の英国滞在中(1987年)、ボブ・シモンズ博士(左端)のお宅でご家族と一緒に。(本人:右端)
カルシウム・シグナルを見る
東北大学の助手として帰国し、骨格筋で興奮収縮連関の研究を始めました。留学してCa2+研究から少し離れている間に、新しいCa2+濃度測定法が開発されていました。細胞内のCa2+に結合してその濃度を可視化する、蛍光指示薬Quin2の登場です。その開発者ロジャー・チェンロジャー・チェン【Roger Tsien】(1952−2016)アメリカ出身の生化学者。緑色蛍光タンパク質(GFP)の発光のしくみ解明に取り組み、遺伝子操作により様々な蛍光を作り出した。これによって2008年下村脩らとともにノーベル化学賞を受賞している博士のロンドン大学でのセミナーをたまたま私は聞いていました。それまでは筋肉の収縮を指標にCa2+濃度変化を推測していたので、変化を直接見ることができるなんて、まさに夢の試薬だと思いました。しかし、周囲の反応は冷ややかでしたね。色素がCa2+を結合することでCa2+シグナルに影響を与えるのではないかなど、アイデア倒れだというのが大方の予想だったのです。でも私は帰国後に早速試薬を取り寄せました。測定装置は市販されていませんから自作するしかありません。試薬を光らせるためには顕微鏡越しに標本に紫外線を当てる必要があるので、顕微鏡メーカーに透過する波長を問い合わせるところから始まり、回路を組み立て試行錯誤の連続でしたね。2年かけてようやくCa2+濃度変化を自分の眼で見れた時は感激しました。
1984年、その2年前に東大に異動されていた遠藤先生から声をかけていただき東大へと移り、Ca2+研究を続けました。細胞内のCa2+濃度上昇には、細胞外からの流入と、細胞内でCa2+を貯蔵している「Ca2+ストア」からの放出という二つのしくみがあります(図1)。当時、平滑筋細胞のCa2+ストアはまだ謎だらけでした。大学院生の堀内桂輔さんが、平滑筋の収縮の強さをCa2+濃度の指標として実験を進めていましたが、非常に苦労していました。そこで、ここに骨格筋用に開発した自作のQuin2の実験系を取り入れました。でも、平滑筋から得られるシグナルは小さく、測定するとノイズだらけで結果がうまく解析できません。平滑筋ではこの手法は使えないかと諦めかけていた時、ロジャー・チェンが第2世代の色素を発表しました。Quin2の30倍信号が大きいFura-2です。日本ではまだ販売されていなかったので、自分で輸入しました。これで見事に綺麗なデータがとれるようになったのです。堀内君と二人で実験結果を見た時、嬉しくてそれまでの苦労が一気に吹き飛びました。いよいよ本格的な実験の開始です。ところが、この実験系では、10種類ほどの溶液を決まった時間間隔で次々入れ替えなければなりません。ちょっとでも間違えるとデータになりません。煩雑な作業が新たな悩みの種となりました。そこで電子回路を実験装置に組み込み、コンピューターを使って全自動化しました。少年時代からの工作と工学部と留学先での学びが実を結び、これを基盤にCa2+放出のしくみを探る私の研究が展開していきました。

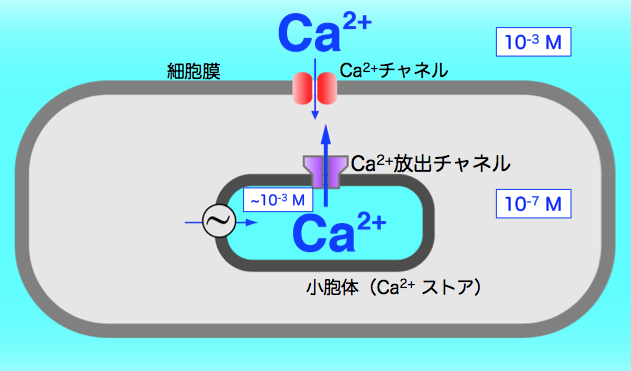
図1:通常、細胞内のCa2+濃度は細胞外に比べて1万分の1程度と非常に低く保たれている。刺激を受け濃度が変化することで様々な機能のスイッチとなる。

東大での最初の10年は階段教室の下の屋根裏部屋のような場が私たちの実験室NC107だった。不本意だったがハングリー精神が芽生え研究の活力になった。

自作実験装置の前で。ここから研究が大きく展開していった。

NC107で実験する大学院生だった廣瀬謙造さん。不規則な形の部屋だったが、効率的に実験ができるよう工夫した。
世界でまだ誰も見つけていない現象をつかむ
1983年、膵臓の外分泌細胞で細胞内Ca2+ストアからCa2+を放出させる作用がイノシトール3リン酸(IP3)にあることがマイケル・ベリッジ(Michael Berridge)博士らによって発見されました。そこで、平滑筋細胞でもIP3に同じ作用があるのではないかと考え、実験を進めました。しかし、いくら平滑筋細胞のCa2+ストアにIP3をかけてもCa2+放出が観察できず、平滑筋にはこの機構はないと結論づけようとしていた時です。ある日、実験手順の入力を間違えてIP3の溶液をいつもとは違う条件で平滑筋にかけしまいました。するとそれまで見たことのない劇的なカルシウム濃度の変化が見えたのです。調べてみると、その溶液にはわずかにCa2+が含まれていました。IP3によるCa2+放出にはCa2+が必要だということです。これは、ストアからの放出によるCa2+の濃度上昇が、さらにCa2+放出を促進する「自己再生産的」な性質をもつことを示しています。こんなに重要なことはもう誰かが見つけているのではないかと図書館の本で片っ端から調べました(まだインターネットはなかったのです)。世界初の発見だとわかった時の気持ちは忘れられませんね。研究人生の中で一番興奮した瞬間です。
自分が見つけた細胞内の現象が、からだの中でどんな意味をもつのだろうと考えをめぐらせました。当時はまだ助手でしたから、研究者としての存在感を示したいと意気込んでいました。IP3によるCa2+放出はストアの膜上にある多数のIP3受容体が制御しています。当時はIP3受容体の実体はわかっていませんでしたが、自分の実験結果からCa2+感受性があることは明らかです。IP3 と微量のCa2+により一つの受容体が活性化されCa2+の放出が始まると近くの受容体が次々に活性化され、ドミノ倒し的に一気に濃度上昇が起こると推測できます(図2)。これは神経細胞で活動電位(興奮)が発生するしくみと似ています。活動電位は細胞膜の多数のイオンチャネルによって制御されており、刺激が一定以上になると興奮し、それ以下では全く興奮しない全か無か(All or None)の反応を示します。ここで平滑筋も全か無かの反応をするのではないかとひらめきました。平滑筋組織内の多数の細胞は一つのユニットを形成しており、すべての細胞が同じように収縮しているというのが当時の常識でした。私は学会で、今までの常識は間違っていると自説を発表しました。例えば、組織全体が50%の力で収縮している時、個々の細胞は全か無かの反応をしており、収縮している細胞が全体の50%、残りは休んでいるという説です。発表が終わるや否や大御所の先生がスクッと立ち上がり「そんなことがあるはずはない。だいたい見てもいないのに、見てきたように言うな!」と厳しい言葉を投げかけられました。
見てもいないのに大きなことを言っても納得してもらえないのは当たり前ですから、早速見るための実験を考え始めました。組織の中での個々の細胞の反応を見るために、動脈の平滑筋に着目しました。動脈は血液と触れる内皮細胞の外側に平滑筋細胞と交感神経が重なっています。交感神経の刺激に応じて、平滑筋の収縮度合いが変わり血圧が制御されています。私たちは血管の構造を保ったまま取り出し、交感神経に刺激をあたえながらCa2+濃度変化と収縮の様子を観察する実験系の立ち上げを目指しました。血管が丸いままだと顕微鏡で観察が難しいため、四角いガラス管を挿入するなど観察の工夫に数ヶ月間悪戦苦闘しました。この実験は不可能かもしれないと弱気になっている頃にようやく成功し、頭の中で予想していた通りの結果が見えてきました。Ca2+濃度は組織内の細胞間で同調せず、個々の細胞が独立の周期で上昇し、収縮について全か無かの反応を引き起こしていると証明できたのです。また細胞内で濃度上昇が徐々に伝播する「Ca2+ウェーブ(波)」や、濃度の上昇と下降をくり返す「Ca2+オシレーション(振動)」も観察できました。Ca2+放出後にストア内がほぼ空になり、再度ストアにCa2+を充填してから次の放出が起きるまでには一定の時間が必要なので、周期が生まれ振動するのでしょう。日本庭園にある鹿威し(シシオドシ)のようなしくみだと私は考えています。一連の結果を学会で発表すると、前年に厳しい指摘をうけた先生にも自説を認めてもらえました。学会の後、ホテルのバーに誘ってもらい、一緒に乾杯してくださったのがいい思い出です。
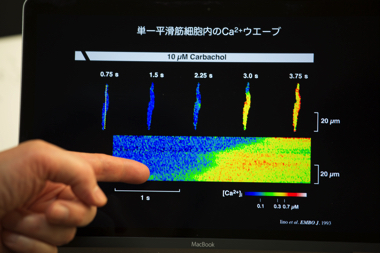
細胞内のCa2+可視化技術によって、濃度上昇の波が見えた。Iino et al. EMBO J. 1993 に掲載した成果。

1991年に来日したハックスリー博士と東大構内にて。左からハックスリー博士、遠藤先生、A.M.ゴードン博士、本人。

マイケル・ベリッジ博士(左)は私が発見した「自己再生産的Ca2放出」を非常に気に入ってくれた。彼のためにベルギーで開催されたシンポジウムに招かれた時の一枚。
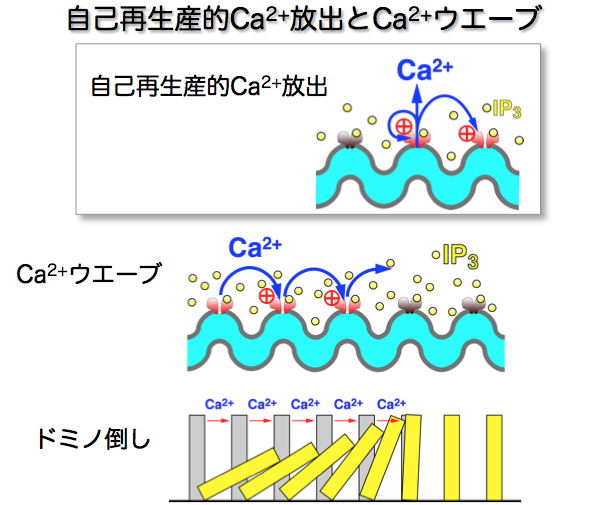
図2:自己再生産的Ca2+放出はIP3受容体を次々活性化し、ドミノ倒しのようにCa2+濃度上昇を引き起こす。
多様な信号が生まれる源
その後、御子柴克彦先生(現・理研BSIシニアチームリーダー) のグループがIP3受容体遺伝子のクローニングに成功し、約2700個のアミノ酸配列(タンパク質の一次構造)が明らかになりました。この構造のどこかにCa2+を感じる部分があるはずだと考え、アミノ末端から2100番目に位置するグルタミン酸が重要な部位であることを見出しました。このグルタミン酸をアスパラギン酸に変えた変異体の細胞は、IP3感受性は正常で、Ca2+感受性だけが低下するのです。細胞内のCa2+濃度変化のパターンを観察すると、一度濃度上昇が起きてもすぐに静まってしまいます。自己再生産的なCa2+放出がオシレーションなど多様な信号パターンを生み出す基本機構であることを実証できたのです。
血管周囲の平滑筋細胞はCa2+オシレーションの頻度によって収縮の度合いが調整されていることがわかり、私たちの研究はCa2+拮抗薬という高血圧の治療薬の作用の理解にもつながりました。現在では、Ca2+オシレーションは多様な細胞のスイッチとしてはたらくことがわかっています。例えば、臓器移植の拒絶反応とも関係します。拒絶反応は移植片を異物(非自己)と認識した免疫系のT細胞の指令により起こります。異物を認識した時、T細胞の中ではCa2+オシレーションが起き複数の遺伝子発現が活性化されるのです。医療の現場では、T細胞内のこの伝達経路を抑える薬が免疫抑制剤として使われています。
私たちはCa2+オシレーションと遺伝子発現の関わりを詳しく追いました。注目したのは、Ca2+濃度上昇によって脱リン酸化されて細胞質から核へと移行し、遺伝子を活性化する転写因子NFATのはたらきです。3分おきのCa2+オシレーションであれば、常にCa2+濃度が高い状態と近い割合で核への移行が起きることがわかりました。短い間だけCa2+濃度を上げてすぐ切ってしまっても、NFATは約7分間Ca2+濃度が上昇したことを記憶しているので、細胞内の反応はCa2+濃度が低くても進むことがわかりました。つまり1回の短い刺激で反応が長続きし、効率がいいのです。Ca2+濃度の上昇は長時間続くと細胞死を引き起こすこともあるので、Ca2+オシレーションは細胞内で重要かつ効率的な伝達方法といえます。

薬理学教室の教授となり、江橋先生がかつて使っていた部屋へ引っ越した。

最初の研究室旅行。助教授の竹島浩さん(前列左から2番目)が分子生物学の手法を導入してくれたことが研究の展開に重要だった。(本人:前列右から2番目)

大学院生の上田泰己さん(後列左から5番目)が当番で企画した2001年の研究室旅行。箱根と小田原に出かけた。(本人:前列左から3番目)
未知機能を探して
遠藤先生との出会いからCa2+シグナル研究へと入り、気がつくと30年以上の月日が経過していました。ある時、医学部の学生との懇親会で「先生はずっとカルシウム研究ですか?」と聞かれました。飽きないのかと不思議なようでした。正直、全く飽きていません。その理由の一つに研究対象の爆発的な広がりがあります。大学院生の頃は筋収縮と分泌ぐらいでしたが、その後Ca2+が発生・分化や免疫だけでなく学習・記憶まで実に多様な機能に関わることが明らかになりました。しかも、今見えているのは氷山の一角でしょう。私たちは2000年ごろから脳に注目してCa2+の未知機能の探索に挑戦しています。研究手法として「Ca2+を見る」ことはもちろんですが、Ca2+は様々な分子と連携してはたらくので、Ca2+以外の情報伝達分子にも着目し、廣瀬謙造助教授(現・東京大学医学部教授)らが中心になって新たな可視化法を開発しました。ここからも研究が新たな展開をしました。
まず注目した分子の一つは、脳内の重要な神経伝達物質であるグルタミン酸です。可視化するために蛍光指示薬を開発し、マウスで運動を司る小脳を観察しました。通常、神経伝達物質は細胞同士の接合部シナプスで伝達されますが、神経細胞に連続刺激を加えたところ、シナプスからグルタミン酸が漏れ出る様子が見えました。現象は知られていましたが、可視化に成功した初めての例です。漏れ出たグルタミン酸はシナプスの外にある代謝型グルタミン酸受容体を活性化し、細胞内でIP3受容体を介したCa2+放出を起こします。そこで、分子生物学的な手法を使ってIP3を急速分解することでこの経路を止めてみました。その結果、IP3受容体を介したCa2+放出が神経栄養因子BDNFを分泌させ、シナプス結合の維持につながっていることがわかりました(図3)。連続刺激によってシナプスから漏れ出たグルタミン酸は、神経間の伝達効率を上げる作用があるのです。自転車に乗るなど運動の記憶は消えませんが、久しぶりだとカンが鈍りますよね。刺激を与え続けることが、能力の維持に大切なのです。それの基盤になっているメカニズムかも知れません。


機能生物学専攻の教授会にて三品昌美先生の退任をお祝いした時の一枚。私が神経系の研究へと移行する時は周囲の先生方の刺激を受けた。左から、本人、三品先生、狩野方伸先生、河西春郎先生、森憲作先生、宮下保司先生。
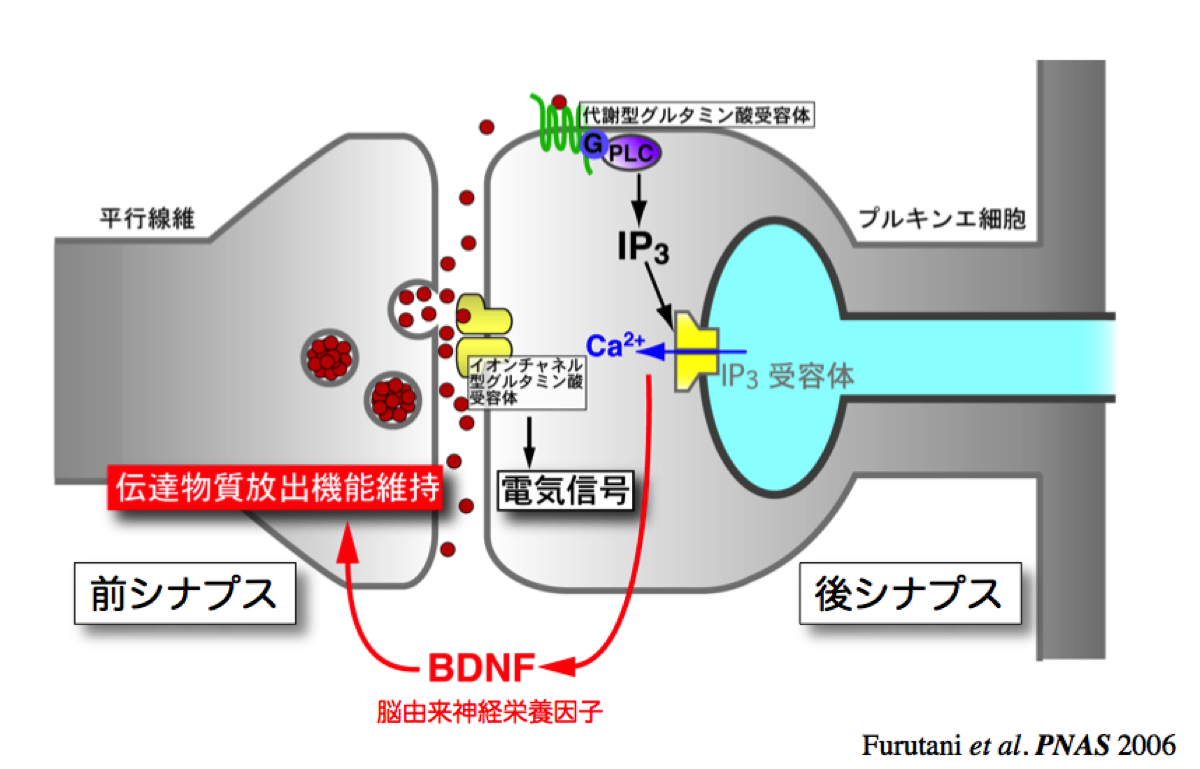
図3:連発刺激によるグルタミン酸漏出はシナプス機能を維持している。
人間の考えることなんてたいしたことない
一酸化窒素(NO)の可視化からもCa2+シグナルの新しい側面が見えてきました。NOは排気ガスなどに含まれる分子ですが、体内では酵素によってアミノ酸から合成され、情報伝達分子としてはたらいています。NO合成酵素がCa2+で活性化されることから、私たちはNOにも興味をもちました。従来、NOは非常に小さい分子なので遠くまで拡散してはたらくシグナルだと考えられていました。100μm以上も移動すると言われていましたが、これが本当なら沢山のシナプスに同時にはたらいてしまいます。可視化技術によって確かめると実は拡散距離は短く、ごく近傍のシナプスにはたらくシグナルだということがわかりました。さらに、神経細胞内でNOがCa2+ストア上のリアノジン受容体を介してCa2+放出を引き起こすことも明らかにしました(図4)。リアノジン受容体はIP3受容体と相同なタンパク質で、一つのタンパク質が二通りに進化してきたと考えられています。
神経や筋肉の興奮にともなって起こる細胞内Ca2+濃度変化は、信号を細胞内に伝える巧妙なしくみです。活動電位は、もし細胞膜の厚さが1 mだとすると、1000万ボルトに達するほど大きなものですが、Ca2+濃度変化に変換されなければこの信号は細胞内部には伝わりません。興奮を伝えるCa2+のはたらきの基本型は細胞膜のCa2+チャネルで、興奮によって開き細胞外からCa2+の流入を引き起こします。心筋では、Ca2+流入によるCa2+濃度上昇がリアノジン受容体を刺激しCa2+放出を起こします。流入によるCa2+の増加をリアノジン受容体が増幅しているのです。また、Ca2+シグナルがより進化した骨格筋では、細胞膜のCa2+チャネルとストア上のリアノジン受容体が隣接しており、Ca2+チャネルの活性化が直接リアノジン受容体を介したCa2放出を引き起します(図4)。からだを動かす骨格筋ならではのしくみです。必要な時に素早く逃げられる方が生存に有利ですよね。興奮とCa2+濃度上昇をつなぐこれらのしくみは実に見事で、リアノジン受容体の機能的な意義が美しく説明できることに常々感心していました。ここにさらに、NOによるCa2+放出という新たな機構があることは、すぐには信じられませんでした。自分の知識が完成していると思い込んでいたのです。助教の柿澤昌さん(現・京都大学准教授)や山澤徳志子さん(現・慈恵医大准教授)らに綿密な実験を重ねてもらい、ようやく結果を受け容れることができました。「人間が考えることなんてたいしたことないんですよ」と遠藤先生がいつもおっしゃっていた言葉が頭の中に響きました。私たちの予想をはるかに超えた広がりと複雑さが細胞の中にはあるのです。この発見の後、NOによるCa2+放出の生理的な意義および病態との関わりを調べています。
最近は、生きたままのマウス脳内のアストロサイトのCa2+濃度を観察する研究も進めています。アストロサイトは神経細胞の活動を助けるグリア細胞の一種です。観察の結果、アストロサイト内で自発的にCa2+シグナルの一過的な上昇が起きていることを見出しました。星のまたたきと重ねて「Ca2+トゥインクル」と名付けました。さらにアストロサイト内ではCa2+濃度の上昇が神経細胞の保護に重要だということが見えてきています。生きている個体内のCa2+濃度変化を観察できるようになるなんて、研究を始めた当初は想像もしなかったことです。急速な技術進歩とともに研究人生を歩めたのは、非常に幸運でしたね。
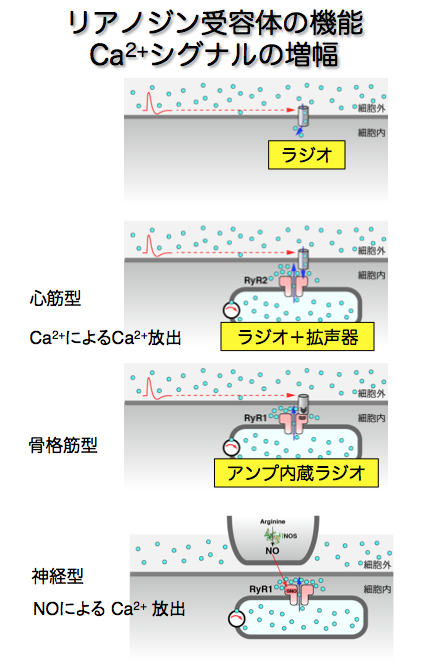
図4:リアノジン受容体はCa2+シグナルの「増幅器」と考えられる。
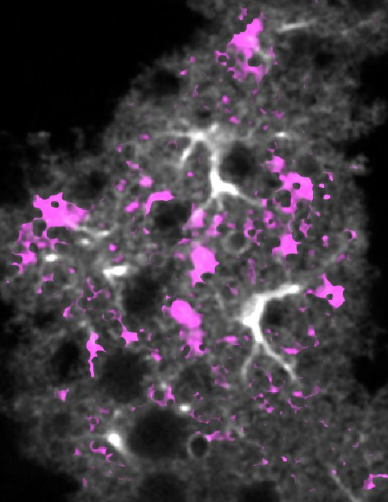
生きているマウスの脳内で見たグリア細胞のCa2+シグナル。微細な濃度変化も逃さず捉えて、詳細に活動を解析できる。
得意なことを伸ばす-放牧主義
研究室のメンバーからは「放牧主義」だと言われます。細かくあれをやりなさい、これをやりなさいとは言ってきませんでした。研究って、実験するとだいたいうまくいかないものです。99%は失敗で、1%ぐらいひょいっとたまにうまくいく。だからこそ好きなこと、自分がやりたいことでないと興味を維持できないと思うのです。それから、メンバーそれぞれに得意、不得意があります。不得意なことを克服することもある程度必要ですけれど、自分のことを振り返ってみても一番得意なところを伸ばせる環境が理想だと思います。だから研究室に入ってきた学生には「何をやってもいいよ、でもカルシウムに関係することをやってね」ってそういう感じです。研究の進展によって、生体内でのCa2+の重要性がどんどん明らかになり、今ではCa2+が何もしていない細胞なんてないのだろうと思っています。これからもCa2+を解析の道具として生きものの機能を探り続けます。


研究室の出身で、東大医学部の教授となった廣瀬謙造さん、上田泰己さんと一緒に3研究室合同のBBQを開催した。

