

普通の子供時代
父は医学部を出て国家公務員になり、戦後に公職追放令で職を追われた人たちの復帰に献身したと聞いています。20万人以上のほとんどが復帰したそうですから、訴願を聞き入れGHQと交渉して追放を解除するのはたいへんな仕事だったのでしょう。過労が祟ったのか僕が4歳のときに亡くなりました。幸い父に恩義を感じた人たちが育英金を募って下さり、僕と妹は目白の母の実家で不自由なく暮らしました。祖父が父親代わりでした。祖父の祖父が会津の白虎隊の生き残り飯沼貞吉の弟で、祖父自身も会津武士の精神をもつ人でした。こう話すとドラマチックですが、平和な苦労のない子供時代でした。将来は、「天文学者になりたい」と言っていましたが、特に理科系が得意というわけでもありませんでした。
中学は質実剛健な進学校、開成に入りました。ところが僕はあまり適応性がないようで1年生のときの成績が下からから数えたほうが早いという状態でした。それに驚いた母が家庭教師をつけたのです。開成を卒業して東大法学部に入学したばかりの学生の新欣樹さんで、後に通産省に入省する秀才でした。この人の影響は大きかったですね。演劇をやっていてその面白さを話すので、中学生の日常とはかけ離れた世界を想像して憧れました。中高生時代で記憶に残るのはそのくらいです。同級生だった山本正幸さん(現・基礎生物学研究所所長)が、普通のかわいい子だったと言っていました。マラソン大会でがんばったり、よく勉強したり、それで十分だったのです。

しっかり者の妹百合子と(左)。3歳と5歳くらい。

カソリック系の目白幼稚園に通った。右端で椅子に座っているのが密かに憧れていた堀さん。(本人:左端、マットに座っている)

父親代わりだった祖父と夏休みに北軽井沢の別荘で薪集め。「お金で買えないものだけが大切なんだ」と常日頃言っていた。(本人:左)

夏休みに北軽井沢の別荘で、新欣樹先生(右)と勉強。新先生は、東大法学部を卒業し通産官僚になった。

館山水泳合宿。とても厳しかったが、中2の時小遠泳で4km泳ぎきり、上がって飲んだ麦茶の味が忘れられない。(本人:中央)

高校2年の開成マラソン。学校の行事も一生懸命がんばった。

高校3年の終わりに5〜6人の仲間と三浦半島一周のサイクリングをした。(本人:左)
演劇青年から科学者へ
母の公務員にさせたいと言う願いを入れ、法学部を目指して東大文科一類を受けましたが、残念ながら不合格。翌年東大の理科二類に入学しました。授業が始まる前に演劇部の部室に向かい、先輩に「明日から朝10時来るように。仕事は5時まである」と言われてから、部室に入り浸りの毎日が始まりました。転落の始まりですね。それでも文科系なら進級できたのかもしれませんが、実験や実習のある理科系ですから、結局、教養部に4年かかってしまいました。でもその頃は、演劇こそが「生きている」という実感を与えてくれるという感じで夢中でした。僕は役者でしたが、10人ほどが集まって脚本を練り、公演が近づくと照明、効果、大道具、小道具、舞台監督と一体となって何日も徹夜して、舞台を仕上げるのです。しかし、何かに憑かれたような熱中の日々に酔いながらも、役者で食べていく才能がないことはわかりましたし、演劇にのめり込んでいる人たちを見てこれではまずいと思い始めたのです。そして、同じように熱中して生きられる職業はなにかと考え、「科学者になろう」と思ったんです。科学の新しい発見を追い求める毎日で、演劇と同じように「生きている」実感が得られるに違いないと思ったのです。生物学者は、子供の頃から昆虫が好きだったというような人が多いと思いますが、僕は24歳の時不純な動機で科学者を職業として意識し、そうなろうと決心したのです。

農芸化学を専攻し、遅れを取り戻そうと好きだった有機化学を一生懸命勉強しました。当時助教授であった森謙治先生のご指導で卒業研究では昆虫のホルモンを十数段階の反応を経て合成しました。有機合成は面白かったのですが、その当時の最先端は人海戦術で複雑な化合物の合成に挑んでおり、ノーベル賞を受賞したウッドワードは100人がかりで100ステップかけてビタミンB12を合成したと話題になっていました。僕は研究者の顔が見えるような独自の発見をしたいと、当時急展開していた生化学を志し、大学院は薬学に決めました。生薬や薬理など薬学系の習っていない科目は本を丸暗記して猛勉強しましたね。幸いに合格でき、国立がんセンター研究所の西村暹先生の研究室に入れていただけました。初めてお会いした西村先生にプロの科学者を感じ、こうなりたいと思ったのが、今につながる出発点です。

大学に入学したての頃。演劇部では役者をやっていた。
tRNAとの出会い
西村先生は、遺伝コドンの解明でノーベル賞を受賞したコラーナの研究室で遺伝暗号の解読をされ、帰国後tRNAの研究を始めたところでした。日本で数少ない一流の分子生物学研究室のひとつで、他の学部からも人が集まっており、渡辺公綱さん(現・東京大学名誉教授)、大隅良典さん(現・東京工業大学特任教授)、横山茂之さん(現・理化学研究所上席研究員)などもいました。故人になってしまいましたが田矢洋一さん(前・国立がんセンター研究所部長)には実験を教わりました。彼は秀才の誉れ高く強い影響を受けました。僕は1年浪人、2年留年しているので遅れている上に分子生物学は初めてですから、懸命でしたが、全く歯が立たず焦りました。修士過程のtRNAの研究では思うような結果が出ず、研究者としての能力に疑問をもち、博士課程への進学はかなり悩みました。博士課程の最初の一年間は生化学部の杉村隆部長のところで、がん組織で変化するアルドラーゼのmRNAの単離をするつもりで、アルドラーゼの精製をしていました。発がんに伴いどのように遺伝子の発現がスイッチされるのかを見ようとしたのです。アルドラーゼは精製しましたが、 mRNA研究は進展のないまま西村研のtRNAの仕事に戻り、杉村先生の研究室には1年お世話になっただけでしたが、自由な発想から多いに刺激を受けました。その後の杉村先生の「おこげの研究とその発展」や「チョウの研究からアポトーシス」などのユニークな研究のアイディアの源泉を理解できたのはよい経験だったと思います。
博士論文は「tRNA中に存在する微量塩基性分Qの生合成機構」。博士1年の終わりに西村研究室で始めた研究が急展開した結果です。何をやっても上手くいかずへこんでいたある日、西村先生が「この論文は面白いから追試をしてみたらどうか」と言われたのです。アメリカのファーカスという研究者の論文で、網状赤血球とグアニン残基を放射能で標識したGTPを混ぜるとtRNAに放射能が取り込まれるという実験でした。西村研究室では大腸菌のtRNAを全て独自の技術で分離精製していましたので、それを使えたのです。鶏の血液からとった網状赤血球の酵素で大腸菌のtRNAにグアニンを取り込ませる実験です。するとヒスチジン、チロシン、アスパラギン、アスパラギン酸の4種のtRNAだけがグアニンを取り込んだのです(図1)。
RNAは通常A、U、G、Cの4種の塩基から構成されていますが、tRNAには特殊な修飾のついた微量塩基がたくさんあります。西村先生はそれがtRNAのコドン認識の特異性に関わると考えて、精力的に研究を進めていました。中でも最も複雑な修飾がQという塩基で、グアニンの環状の7位のNがCに置き換わったデアザ構造を持ち、西村先生が苦労してその構造を決めました(図1)。僕が見つけた4つのtRNAは、すべてアンチコドンの1文字目にQの修飾をもつtRNAだったのです。そしてグアニンの取り込まれたのが、まさにQの位置だということを証明できました。鶏の網状赤血球中にある酵素が、大腸菌のtRNAのアンチコドンの1文字目のQ塩基をグアニン塩基にすげ替えたのです(図2)。これは、tRNAの修飾反応で初めての発見です。Q塩基の生合成解明の糸口をつかんだと思い、次の作戦を練りました。デアザ構造を持つQの前駆体の塩基が単体で合成され、それが塩基置換によってtRNAに取り込まれ、さらに修飾反応を受けてQに成熟するのではないかと推理し、その全体像を確かめようと決めました。朝8時半から深夜2時頃まで実験する生活を丸2年ほど続け、Q塩基の生合成経路を決めました。これで、生化学会奨励賞を受賞しました。tRNAの微量塩基が塩基の置換反応で合成される例はこれだけで、この分野の重要な発見として今でも引用されています。

この経験が僕の研究のスタイルを決めたと思います。最初の発見を嘘かもしれないと「疑い」つつ、でも本当ならとても面白いはずだとエキサイトしながら、何ヶ月もかかって証明するやりかたです。この「疑う」期間が長ければ長いほど研究は独創的なものなのになるではないかと思います。ゼロから始め、それを自分の手でストーリーを構築することが研究の醍醐味で、そうでなければ研究ではないとこの経験から思っていました。
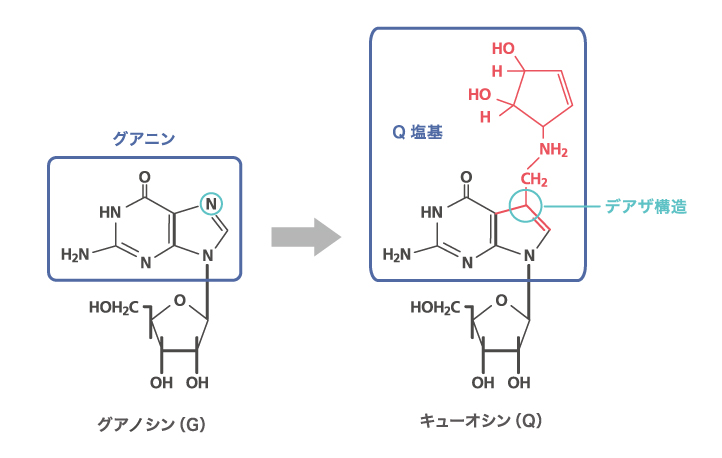
図1:tRNAのグアニン塩基とQ塩基の塩基置換が関わる反応で、Q塩基が生合成されることを発見した。
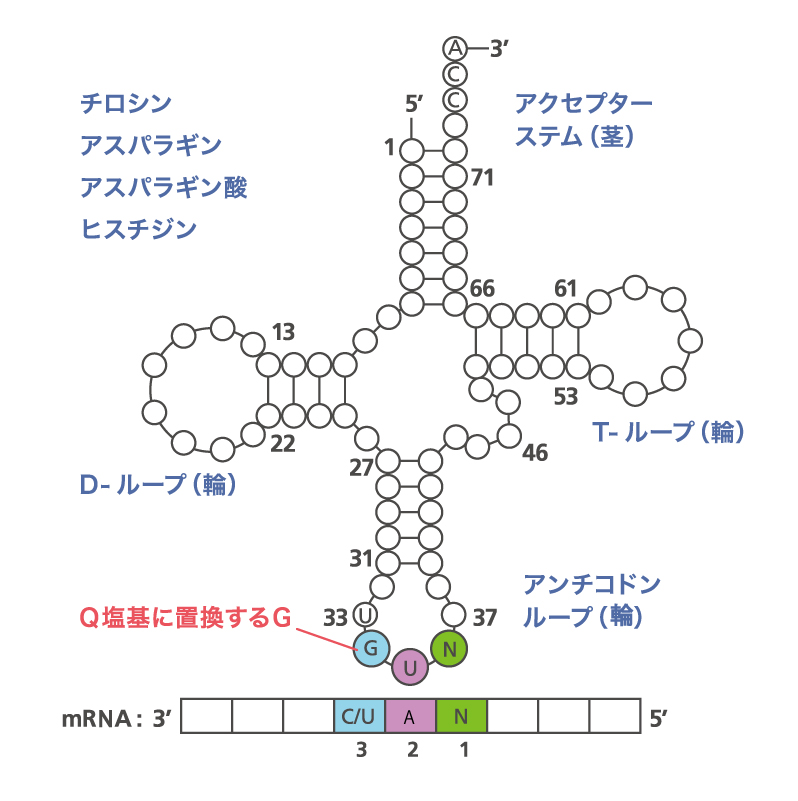
図2:tRNAの構造。クローバーの葉に似ているのでクローバーリーフ構造と呼ばれる。
反復配列の発見
学位取得後10ヶ月アメリカのNIHの畑中正一先生の研究室のポスドクをしていたのですが、筑波大学から講師の誘いがあり帰国しました。32歳のときです。分子生物学研究室には大島靖美さん(現・九州大学名誉教授)が助教授でおられましたが、講師でも独立して研究ができる環境でしたので、独自の研究を始めようと意気込みました。tRNAでは西村先生と競争してもかなわないと思い、やめる決心をしていたので、得意なRNA分析技術となにか最新の技術を結びつけてできることを考えていました。試験管内でDNAからRNAへ転写する技術に出会い、全ゲノムDNAを試験管内で転写して、読まれたRNAを電気泳動で分離しようと考えました。この実験を思いついたのは当時の池村淑道さん(現・長浜バイオ大学名誉教授)の研究の影響です。彼は大腸菌の全tRNAを二次元ゲル電気泳動で分離するシステムを構築していました。今はゲノム研究のおかげで全体を見るという研究が主流になっていますが、1980年代当時は現象の全体を見るという発想はなく、池村先生が初めて生物学の分野にこのような発想を導入したのだと思います。それで、ゲノムから転写されたRNA分子全部を電気泳動で分けるとどう見えるか、ゲノム中に存在する全プロモータープロモータDNAからRNAへの転写の際、RNAポリメラーゼに認識され、転写を開始するシグナルをもつ配列。をマップ出来ないか、とにかくやってみることにしました。
マウス全DNAの転写産物の一次元電気泳動は200塩基くらいの位置にバンドが見えました。つまりマウスのゲノムには200塩基ぐらいの同じ遺伝子がたくさんあるということです。これは面白いと予感し、さまざまな生物で比べてみようと思いました。研究室が解剖室のようになる中で、転写をしてみると、イモリ、カメ、シロサケで明確なバンドが1本見えたのです(図3)。しかも、長さは生きものごとに違う。頭を捻りましたね。タンパク質遺伝子は遺伝子ごとに長さが異なるし、コピー数も少ないのでこんな風には見えないはずです。tRNA遺伝子は数百コピーあるはずですが、泳動の位置からしてtRNAではありません。数千数万という似た配列を持つ非常にコピー数の多い分子がRNAポリメラーゼのプロモーターを持ちそこから転写されていて、この方法で検出されるとしか考えられません。この塩基配列を知りたいわけですが、当時は500塩基決めるにも1年くらいかかりましたから、狙いを定める必要があります。サケで鮮明なRNAのバンドが6SS値S値は沈降係数のこと。分子の大きさを測る目的で、平衡密度勾配遠心法で分子を分離した時の単位加速度あたりの沈降速度。RNA分子では150-200ヌクレオチド前後。 に近い位置に出るので、その塩基組成を分析したところ、A、U、G、C以外にシュードウリジンが出てきたのです。これはtRNAに含まれる微量塩基成分で、学生のときからtRNAを見慣れていたのですぐにそれとわかりました。つまりサケのゲノムDNAにコードされ、RNAポリメラーゼで転写される反復配列にtRNA様構造があるという可能性があるというので興奮しました。おそらく核抽出液に含まれるtRNAの修飾酵素がそのtRNAに似た配列を認識して、シュードウリジンができたのでしょう。大学院生の松本健一君(現・島根大学教授)が1年くらいかけてその配列を決めたところ、みごとにリジンのtRNAに似た構造があらわれました(図4)。シロサケのゲノムにある反復配列がリジンtRNAを起源としてできたことを発見したのです。
もう一人の学生の坂本和一君(現・筑波大学准教授)が、他の動物の既に報告のある反復配列にtRNAと相同な配列があるかを調べました。すると、霊長類のガラゴ、ウシ、マウス、ウサギと調べた全ての反復配列からtRNA様配列が見つかりました。tRNAはゲノム中にほぼ50種類もあり一つ一つ配列も異なるので、それらと反復配列との相同性を調べた人はいなかったのです。tRNAを長い間研究してきたからこその発見でした。
ところが論文を出したら受理されないのです。大発見と勇んでNatureに出しましたがダメ、PNAS(米国アカデミー紀要)もEMBOジャーナルもダメでした。そこで木村資生先生に相談したら、Journal of Molecular Evolutionを紹介して下さったので、坂本君の比較の結果を投稿し、ようやく受理されました。たまたま訪れたエール大学で、そこの教授(アラン・ワイナー、のちに友人になる(写真5-1)が、同じ結果の論文がNatureに受理されたと言うのです。その論文の投稿者は実は以前論文の別刷を送った相手で、自らライバルにヒントを与えてしまったとわかり、心底がっかりしました。まだ30代、駆け出しの日本人ですから世界での駆け引きを知らなかったのです。でも負けていられません。木村先生に相談すると、ポスドクの遠藤浩君(現・金沢大学教授)のイモリとカメの反復配列がtRNA起源であるという論文をPNASに推薦してくださいました。日本学士院紀要にも働きかけてくださり一番早く論文になりました。
サケの反復配列にリジンのtRNA構造があり、転写のときにシュードウリジンが合成されるという松本君の論文は、エームズ試験で著名なカリフォルニア大学のブルース・エイムス教授にPNASの推薦を頼みました。彼はtRNAのシュードウリジン合成酵素がヒスチジンのオペロンの発現に関係しているという面白い研究をしていて、興味をもってくれると考えたのです。面識もないのに不躾なお願いだったのですが、喜んでPNASに推薦してくれました。ライバルのNatureでの論文発表の前後に我々の4本の論文が受理されました。その論文をNatureに送ったところライバルの論文掲載のとき解説記事に引用してくれました。今ではtRNA起源のSINE(短い散在性反復配列、レトロポゾンレトロポゾンゲノム内を移動する転移因子「トランスポゾン」のうち、自分の配列をRNAに転写した配列から逆転写酵素によりDNAに複写された配列をゲノムに再挿入して増幅するもの。の一種)の発見者としての僕のクレジットを多くの学者が認めてくれています。支援して下さった木村先生には本当に感謝しています。

まだゲノム配列もあまり読まれていない時代に反復配列を見つけ、それがtRNA起源だと証明したのは大きな発見だと思いますが、新しい発見をするということとそれを世界に認めさせるということは全く次元の異なることだと痛切に思い知らされました。これを教訓に、第一発見者として追従を許さないよう徹底的にやると思い決めました。まず、tRNA起源のSINEが無脊椎動物、植物、カビと真核多細胞生物の調べたほとんどの種に存在することを示しました。そこから「SINEどうは増えるのか」、「それが種や生物のグループの系統とどう関わるのか」、「SINEにはどんな意味、機能があるのか」という疑問が生まれました。ここから生涯をかけてこの疑問に取り組むことになります。

40歳の頃健康診断の数値が悪く、運動のためエアロビクスに挑戦。3ヶ月で数値は正常になり、いい気分転換にもなった。
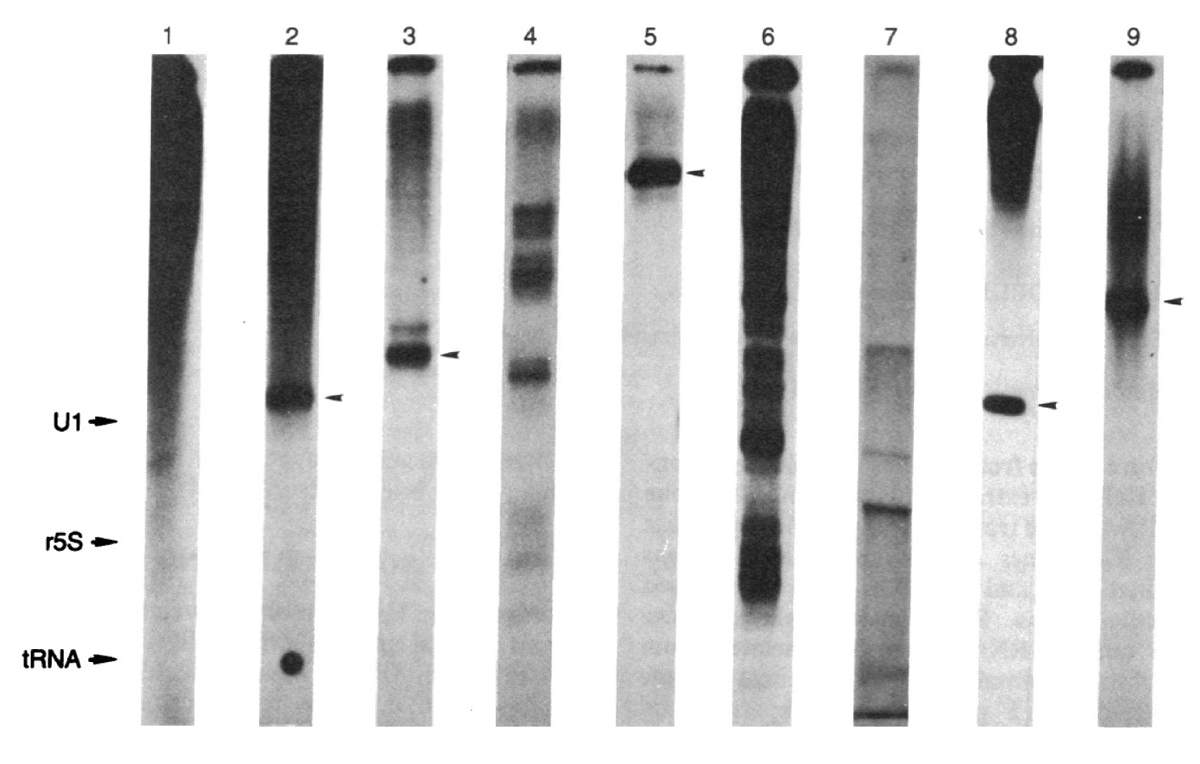
図3:ゲノムDNAを試験管内で転写させた時にはっきりとしたバンドが現れる。のちにこれがSINEからの転写産物であることが証明される。1:ヒト、2:マウス、3:カメ、4:カエル、5:イモリ、6:肺魚、7:シーラカンス、8:シロサケ、9:ヤツメウナギ
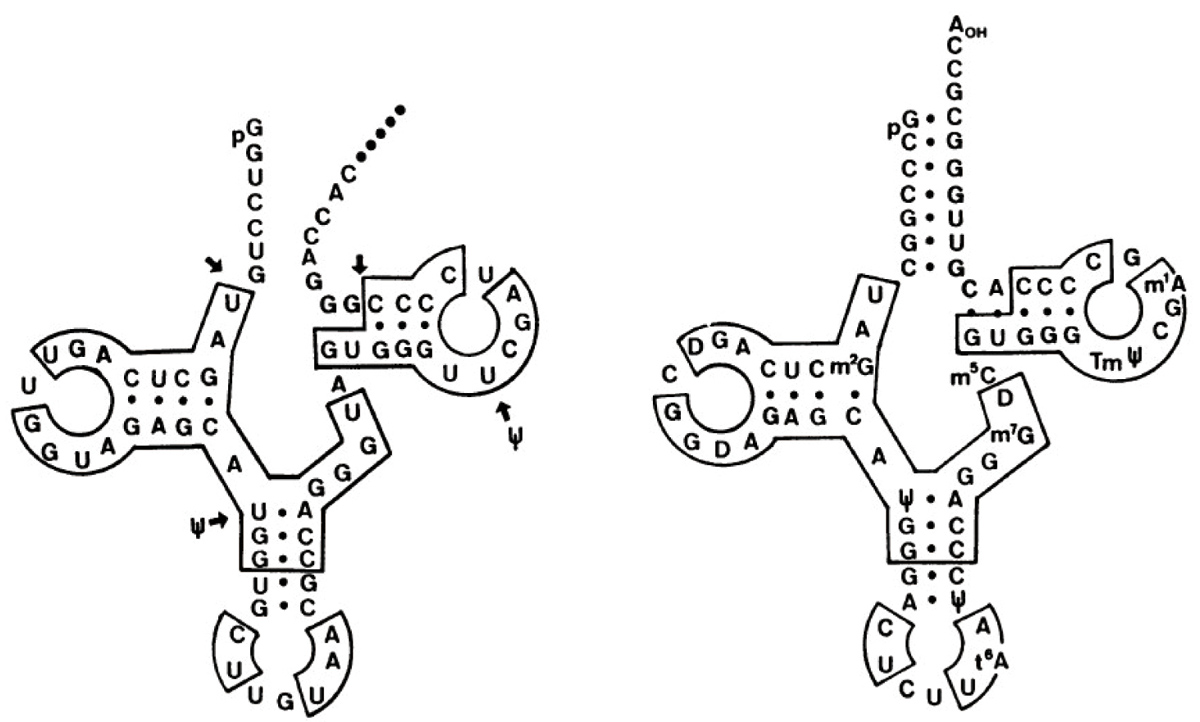
図4:シロサケの6S RNA配列から見つかったtRNA様構造(左)。右はその起源と考えられるリジンtRNA。
SINEはどうやって増えるのか
tRNA起源のSINEは、tRNA様の配列以外の部分は種によって長さも配列も異なりタンパク質もコードしていないのに、なぜゲノム中で増幅することができるのか、それが第一の疑問です。膨大な数存在するのですから、そのメカニズムを知りたかったのです。これは、SINEとは別なレトロポソンのLINE(long interspersed elements、長い散在配列)との関係が鍵となりました(図5)。大島一彦君(現・長浜バイオ大学准教授)がカメのSINEはLINEと3’末端の配列が似ていることを見つけました。SINEと同じようにLINEもまずRNAに転写され、それがDNAに逆転写されてゲノムに入るのですが、LINEには逆転写酵素がコードされていて自分で転移できます。LINEの逆転写酵素はLINEの3’末端の配列を認識することがわかっていたので、SINEの3’ 末端にLINEと似た配列があるということは、LINEがコードする逆転写酵素がSINEも認識し、同じメカニズムで転移すると推測できます。3’ 末端に相同性をもつSINEとLINEがペアになっており、LINEの逆転写酵素によってSINEが増幅するというモデルができました(図6)。そこで、同じようなSINEとLINEのペアが他にも見つかるか、調べたところSINE はどれもある特定のLINEと末端を共有していました。このメカニズムを実験的に証明したのが梶川正樹君(現・東京工業大学講師)です。ウナギのSINEとLINEを用い、SINEの3’末端の配列が似た配列を持つLINEにコードされている逆転写酵素によって厳密に認識され、転移されることを証明しました。認識される配列は、ステム(茎)アンドループ(輪)という二次構造をしており、ループ構造がこの「厳密な認識」には重要でした。この論文はCellに出版されています。一方で、tRNA様構造をもたないヒトのSINE のAlu配列の増幅に関わっているL1というLINEの酵素は厳密に配列を認識していない、「緩い認識」だとされています。この違いも解き明かそうと増幅のメカニズムの仕事はまだ進行中です。
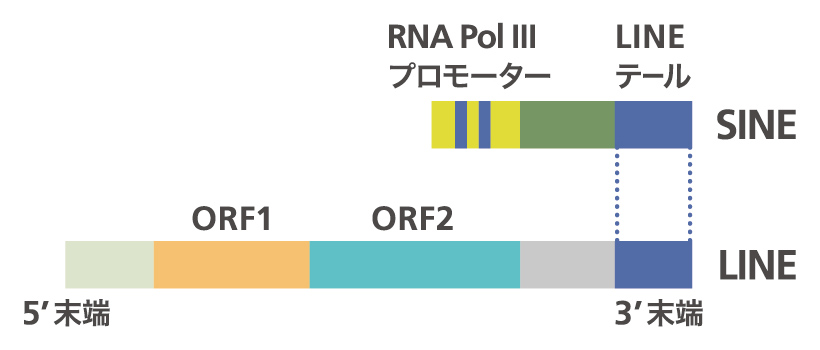
図5:SINE(Short INterspersed Elements)とLINE(Long INterspersed Elements)。SINEは、5’側にtRNAや5S RNAなどのRNAポリメラーゼIIIで転写される配列があり、3’末端はLINEの3’末端と似た配列がある。LINEは、逆転写酵素など転移に必要な酵素をコードしている。
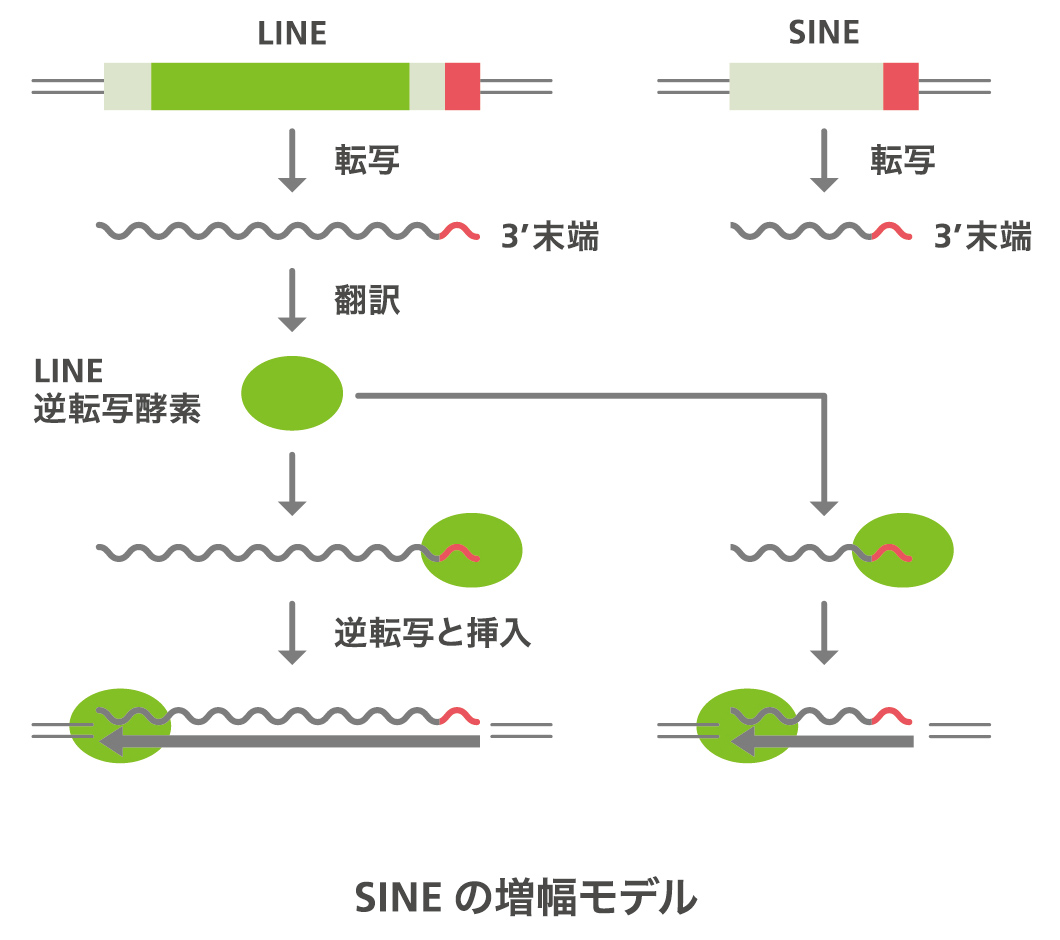
図6:SINEはLINEの逆転写酵素に3’末端を認識され、ゲノム中に挿入される。

写真5-1:1994年12月神戸の分子生物学会に呼んだアラン・ワイナーと三宮で食事をした。翌1月阪神淡路大震災がおき、アランから日本で出会った人々の無事を祈る悲痛なメールをもらった。
サケから始めたSINE法
SINEが殆ど全ての多細胞真核生物ゲノム中に広がっているという、大きな描像が描けたので、レトロポゾンがゲノムの成り立ち、即ち種の形成にどう関わるのかということを考えました。レトロポゾンは一度DNAからRNAに転写され、それが逆転写されたDNAがゲノムに再挿入されるコピーアンドペースト方式で増えます。つまり一度入った配列はもう移動しないので、ゲノムに痕跡を残すのです。そのようなSINEの存在がゲノムの歴史を語らないはずがありません。それをサケで確かめました。
まずSINE配列が、サケ科全体にどのように分布しているかを調べました。サケ科はサケ属、サルモ属、イワナ属など11属あるのですが、我々が最初に見つけたシロサケのSINE配列は、同じサケ属の近縁種カラフトマスにしかないことがわかりました。他にイワナ属にだけある配列、サケ科全体に分布する配列も見つかりました。これを総合するとSINEが異なる系統で異なる時期にゲノムの中で増幅すると推測されます。新しいSINEファミリーは生物のある系統で突然現れ、増幅を始めてその系統の生物の中で広がるという性質をもつのです。
ここで、サケ科に分布するSINEがいつどのように増幅し、どのように分布しているかを調べることにしました。SINEの配列の両端のプライマーを作成し、それがサケ科の他のサカナでも同じ座位座位ゲノム中の位置のこと。特定の1カ所を指すことも、同じ種類の配列の位置をまとめて指す場合もある。遺伝子の場合は、遺伝子座となる。に入っているかを見ることで、そのSINEがいつどの系統で入ったのかが分かります(図7)。このSINEの挿入の有無を指標にして系統樹を作れるのではないかと思いつきました。そこでサケ属のSINEを含む座位をPCRで増幅してSINE座位を探したのですが、1991年当時はこれはたいへんな仕事でした。座位を見つけて、配列を決めてPCRをするのですが、配列をとった種から離れた種では、配列があっても変異が多く検出できなくなるので、よく保存されている配列を探さなくてはなりません。大学院生の村田成範君(現・武庫川女子大学准教授)が3年がかりでサケ属の系統を決めました。結論は、シロサケとカラフトマスが単系統、その外にベニザケがいて、それがまた単系統でマスノスケとギンザケが単系統で前の3種のクレードと姉妹群姉妹群祖先が同じ2つの単系統群をお互いに姉妹群と呼ぶ。をつくり、その外にサクラマスがいます。サケ属の系統はこれが決定版です。これがSINEの有無を指標にして系統樹をつくるSINE法を報告した最初の論文になりました。
最初のSINEを単離したのがシロサケだったので、サケ科を用いて基本的なことが一通りわかりとても幸運だったと思っています。ただ、普遍的な生命現象を提案するにはサケではインパクトが弱いので、生物学でもっと注目されている生物に目を向けようと決めました。

ミュンヘンの学会でのパーティ。アトラクションで、誰が一番長くジョッキを持って手を延ばしたままで耐えられるか競争して優勝した。(本人:列左から2番目)

49歳の時に出会った人と再婚し、彼女との新婚旅行。パリの喫茶店で。

ギリシア島での会議に参加したときのスナップ。左手前:池村淑道さん(当時・国立遺伝学研究所教授)、左奥:ヴェルナー・アーバーさん(制限酵素発見で1978年ノーベル賞を受賞)、右手奥:五條堀孝さん(当時・遺伝学研究所教授)。(本人:右手前)
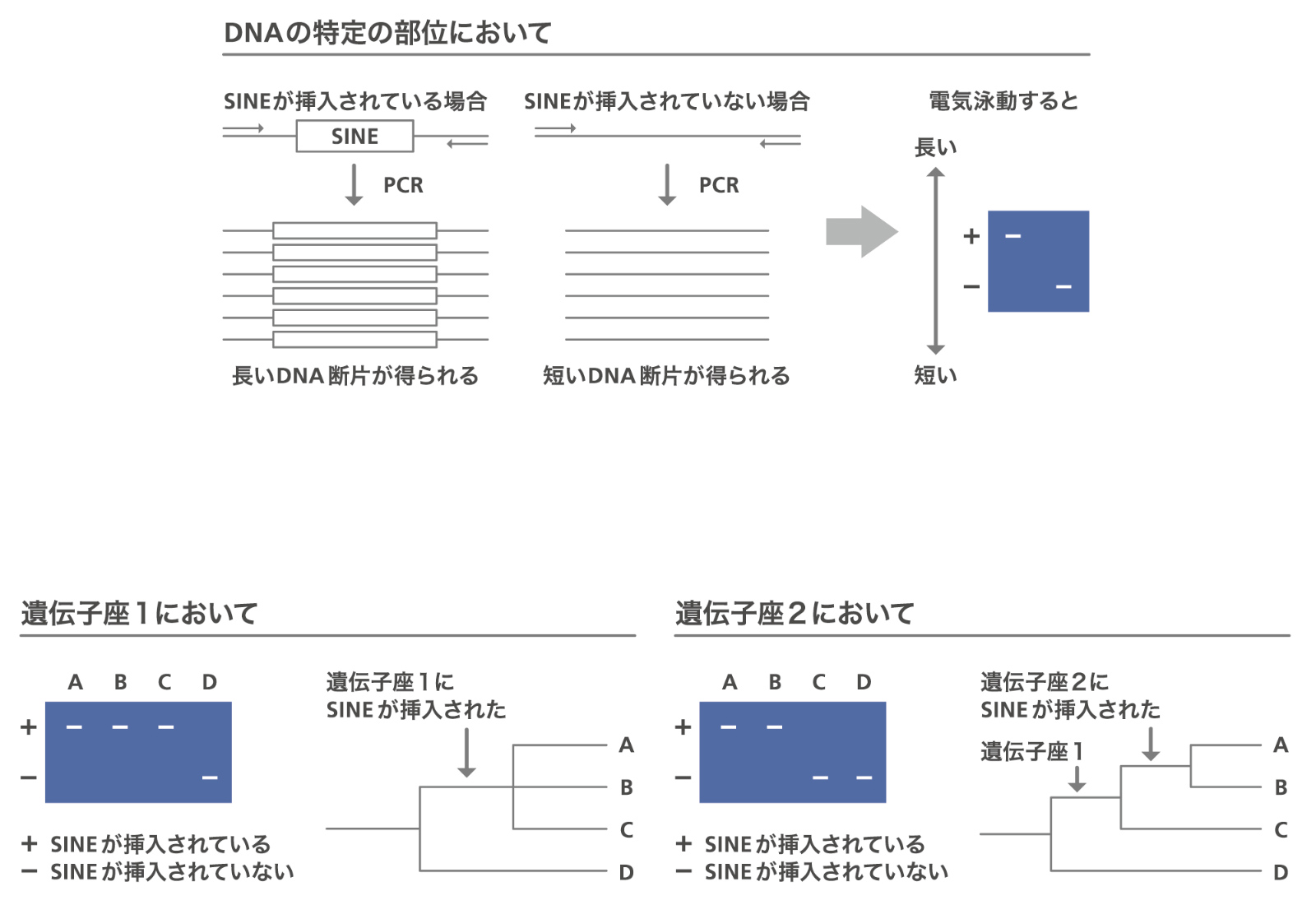
図7:SINE法の原理。同じ位置にSINEが挿入されていれば、PCRで増幅すると電気泳動で長いDNAが検出され、同じ系統に属することがわかる。
クジラから広がる進化
今では僕も哺乳類の系統学の専門家ですが、始めたばかりのころは哺乳類の分類を知らず長谷川政美さん(現・復旦大学教授)にひとつひとつ教わりました。そして、当時論争になっていたクジラの系統解析にSINEで決着をつけることを思いつきました。
クジラの系統は、形態や生態からハクジラ(歯鯨亜目)とヒゲクジラ(髭鯨亜目)に分類されています。ところが分子系統の解析では、この伝統的な系統とは異なる結果が得られ、10年以上にわたる論争となっていました。長谷川さんが論争に加わっていたので、まずそもそもクジラが哺乳類の中でどういう位置づけかという問題に取り組みました。大学院生の島村満君が哺乳類のSINEを調べたらウシとカバとクジラにだけにある配列が見つかったのです。つまり、クジラは偶蹄類の内部系統から進化したことになります。これはクジラの祖先は偶蹄類であることを示します。これは古生物学の説を覆す結果でした。クジラの祖先はハイエナに似たメソニクスという陸上哺乳類であると、化石の歯の形の比較などから言われてきました。メソニクスのくるぶしの骨の形はイヌと似ており、偶蹄類の特徴である滑車状の構造を持っていないのです。言い換えると古生物学者はクジラの祖先は犬に似た動物で偶蹄類とは考えていなかったのです。我々の論文はNatureに載りましたが、世界の形態学者は簡単には認めません。その後ムカシクジラの仲間のパキケタスの化石に滑車状の構造がみつかりクジラが偶蹄類であることが形態学的にも裏付けられ、今では鯨偶蹄目という分類群もできました。
次の問題は、偶蹄類の中での関係です。二階堂雅人君(現・東京工業大学准教授)が、ラクダ、ブタ、シカ、ウシ、カバなどの偶蹄類とクジラ13種から共通するLINEとSINEの座位を探して比較したところ、クジラとカバにだけに共通に挿入されているSINEの座位を発見しました(図8A)。これらの結果よりクジラとカバが最も近縁で、最も遠いのはラクダだと結論しました(図8B)。この研究はSINE法で明らかにされた系統関係の研究としていろいろなところで紹介され、僕の研究で一番有名かもしれません。

次はクジラの系統関係です。マッコウクジラやカワイルカが含まれるハクジラは、歯をもちエコロケーションをします。ヒゲクジラはオキアミなどを髭で漉しとって食べるクジラです。ミリンコビッチは、ミトコンドリアの12Sと16SのrRNAの解析で、ハクジラのマッコウクジラがヒゲクジラに近縁であるという説を出しました。またスウェーデンのアーナソンは、ミトコンドリアチトクロームbの配列から、マッコウクジラがクジラの系統で最も先に分岐し、ヒゲクジラとイルカが近縁だと言いました。形態学や生態学の研究者が見れば荒唐無稽な話です。そこでSINE法でこの問題を解決しようと試みたのです。この頃には解析につかう座位を探すための知識も増え、技術も向上していたので、ハクジラ、ヒゲクジラのそれぞれの系統でゲノムライブラリーからSINEをスクリーニングし、起源が同じSINEが挿入されている座位を確実に単離する技法を確立しました。その結果、ハクジラとイルカが単系統でヒゲクジラが独立の系統であるという、形態による解析を支持する系統樹ができました。もう一つの宿題だったカワイルカの起源も武漢のサンプルを使って中国の研究者と共同研究しました。長谷川さんや二階堂君たちと武漢の水生生物学研究所に泊まり込んだのは懐かしい思い出です。三つの異なる大陸に分布するカワイルカは、異なる川にいるのですが、形態はお互いに似ているので、単系統性が問われていたのです。ガンジス川、揚子江、アマゾン川、ラプラタ川に棲む4グループのうちガンジスカワイルカは、他のカワイルカと単系統を作らず、その形態的特徴は収斂だという結果になりました。そして、クジラ全体でみるとガンジスカワイルカはハクジラの中でも古くに分岐していました。クジラ全体の系統では形態の結果を支持し、カワイルカの系統では形態の結果を支持しないという形で、最終的にSINE法によりクジラ論争に決着をつけることができました(図9)。
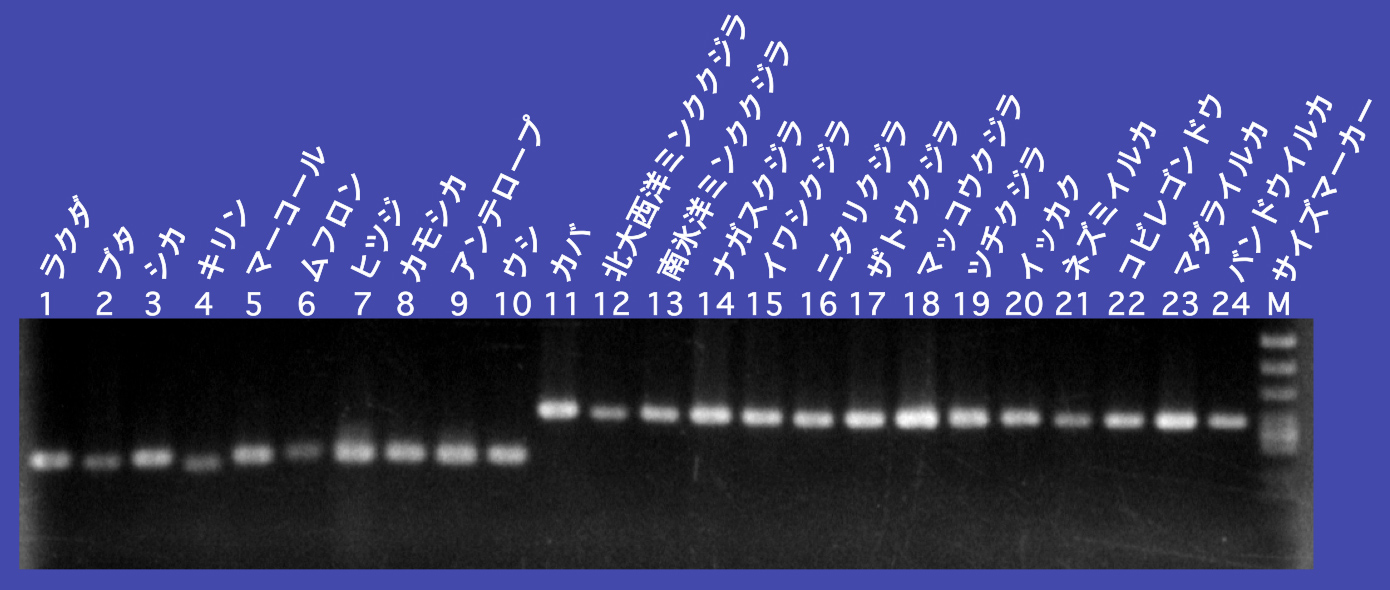
図8A:PCRのデータで示したカバとクジラにだけ見つかるSINE法の例。
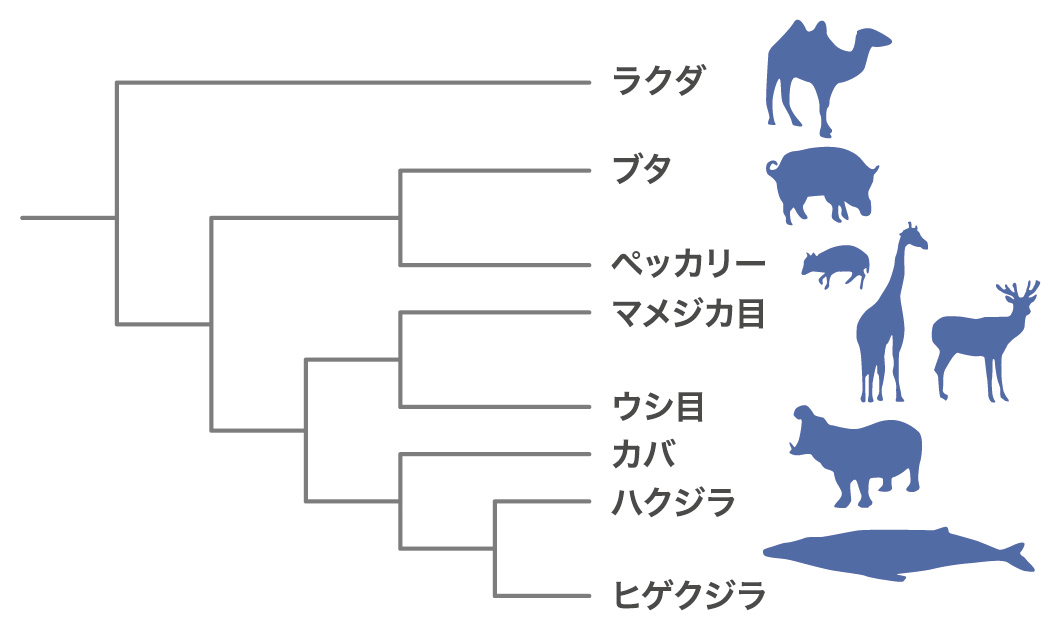
図8B:偶蹄類の中でカバとクジラが近縁であることを示した系統樹。分子進化で一躍注目を浴びた研究。

図9:SINEによるクジラの系統樹。これで論争に決着がついた。

長谷川政美さん(当時・統計数理研究所教授)と曹纓さん(当時・統計数理研究所)とでドイツの学会で。

ジョルジョ・ベルナルディ主催のコスタリカの学会で。左ダン・グラウアー、右ジェルジイ・ユッカ(故人)、後ろはロイ・ブリテン(故人)。(本人:中央)
哺乳類の全体像を描く
クジラの系統解析を通じて、SINE法の威力を証明できました。当時は哺乳類の進化の全体像はほとんど解明されていませんでした。DNA塩基配列の比較による系統解析の全盛期なのですが、配列比較では統計的確率でしか物が言えないのでどうしても多くの間違った報告がありますし、またどうして間違ったかということを検証するのも難しいのです。SINE法で出された結果はホモプラシーホモプラシー共通祖先に由来しない類似(非相同)をいう。塩基配列においては、同じ位置に複数回の置換がおきる(多重置換)などが原因となる。が存在しないのでそれが決定版ということです。SINE座位を探すには経験がいるので僕たちが有利ですが、SINE法が普及して他からの論文も増えました。SINE法がそれだけパワフルだということを示した功績は大きいと思います。
クジラと同じくらい特殊化した哺乳類と言えばコウモリです。コウモリの系統的位置もSINE法で解決しました。かつて形態による分類ではコウモリは霊長類やヒヨケザルと同じグループに分類され、大コウモリ類と小コウモリ類に分けられていましたが、塩基配列の比較から大コウモリがより霊長類に近縁であると言う説が物議をかもしていました。助手の西原秀典君が苦労して4つの異なるSINE座位をみつけ、最終的にはコウモリを奇蹄類や食肉類と同じグループに位置づけました(図10)。そこで長谷川さんと知恵をしぼって、食肉類とセンザンコウのグループferae(フェラエ)に空を飛ぶコウモリと奇蹄類のウマから連想したペガサスを組み合わせて「ペガソフェラエ」という名前をこのグループに対して提案しました。洒落てるでしょ。発表当初はあまり反響がありませんでしたが、最近他の手法でも同じ結果がいくつもでてきたので、ペガソフェラエが定着するのではないかと思っています。

SINE法は非常に確実な方法で、多くの論争に終止符を打つことに貢献しました。しかし、SINEでは決められないケースもわかってきました。種内でSINEの転移があり集団中で多型になっている状態で、非常に短期間の間に次の種分化が起きると、分岐した系統ではSINEが固定するか消失するかは遺伝的浮動により偶然に左右されるので、SINEの有無では分岐の順序がきまらずに、矛盾する座位が現れるのです。(図11)。このような時には系統を決めることはできませんが、SINEの矛盾する座位が得られるということが、種分化が非常に短い時間に連続して起こったことの証明になるのです。このような矛盾座位の存在は、ヒトと類人猿の間でも見られます。SINEの矛盾した座位から種分化が非常に短い間に起きたことを証明できるのだと主張する論文も幾つか書きました。
さらに、このような矛盾するSINEの座位の分析から地質学へと発展した研究があります。哺乳類全体は三つの大きなグループに分けられます。ローラシアに起源を持つ北方獣類、アフリカに起源を持つアフリカ獣類、南米に起源を持つ異節類に分かれますが、その三つのうちどの系統が最初に分岐したかが非常に大きな問題で決まっていませんでした。ところが、レトロポゾンのLINEの配列を使って(SINE法と原理は同じ)分岐順序を決めようとすると、矛盾する座位が多数見つかり三つのグループが短い期間にほぼ同時に分岐したという結論になりました。このデータより、この三つの系統が同時に分かれたのは、ローラシア大陸、アフリカ大陸、南米大陸の三大陸がほぼ同時に分かれたために、大陸間での動物の行き来がなくなったためではないかと考えたのです。そこで大陸移動の専門家である丸山茂徳さん(当時・東工大教授)に、共同研究を申し込みました。地質学者は、まずローラシアとゴンドワナが1億4千万年前に分かれ、1億5百万年前にゴンドワナが南米とアフリカに分かれたと考えていました。この時に彼らが年代測定に用いた地磁気縞は、大陸が海に隔てられた後の海底測定法であり、実際に大陸が離れ動物の交流ができなくなった時期はそれよりも前のはずですが、このような地質学者の用いる手法の理解なしに生物学者はそのデータだけを用いていたのですね。より現実的な膨大な堆積層の掘削データがあると丸山さんに教えられ、アフリカ大陸と南米大陸がいつ分かれたかをひたすら調べました。掘削データから海底に沈んでいる状態と陸に上がった状態の地質を明確に区別できるのです。その結果、ゴンドワナ大陸がアフリカと南米に分かれたのは、1億5百万年前ではなく、1億2千万年前頃に遡るデータが見つかりました。残念ながらゴンドワナ大陸とローラシアが分かれた場所のデータがなく完全な証明にはいたらなかったのですが、三つの大陸はほぼ同時に1億2千万年前に分離し、そのときに地理的隔離によって有胎盤類の共通祖先が三大陸に分断され、それがそれぞれのグループの祖先になっているという新しい仮説を提出しました(図12)。これは掘削データとDNAの結果をもとに新しい地質学の仮説を提示した世界で最初の例でしょう。面白いと思ったことを証明しようとしたら、違う分野の人にも積極的にアプローチすることですね。自分の専門だけやっていたら決してできなかった研究へと展開できたと思います。


図10:哺乳類の系統樹。赤線部分が大陸分断とともに同時に分かれたと考えている哺乳類の三大グループの分岐。

図11:種内に多型がみられる間に短期間で種分岐が連続しておきると、このように矛盾したSINEの分布が検出される。順番が決められないくらい短期間に分岐が起きたことを示している。
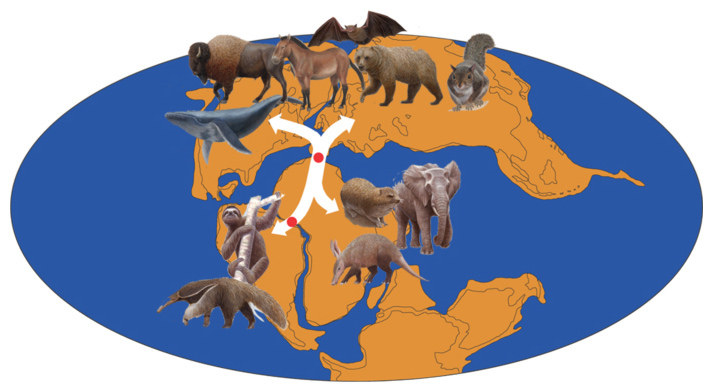
図12:同時に三大大陸が分裂したことにより、哺乳類の三大グループが分岐したという仮説を提案した。(絵:菊谷詩子)

海外の学会は家族同伴の研究者も多いので、妻と娘を連れてベニスの学会へ。

2006年度、第47回の藤原賞に選ばれる。テーマは「生物進化の機構解明」。同時受賞は、村井眞二博士。前列右から4番目:妻彩子、本人、村井眞二博士、村井夫人、後列右から4番目:娘彰子。

2007年の紫綬褒章受章で受賞者と。北大路欣也さん(前中央)。後列左から、本人、藤野陽三さん(当時・東大工学部教授)、妻彩子、藤野夫人。

我が家でのホームパーティ。右手前から、郷通子さん(前・お茶大学長)、長谷川政美さん(前・統計数理研究所教授)、ゾン・ヤン氏(復旦大学教授)、左手前から、妻の彩子、磯崎行雄さん(東大教授)、丸山茂徳さん(前・東工大教授)、中央:本人。

ホームパーティ。右手前から、渡辺公綱さん(前・東大教授)、西村暹先生、左手前から、妻の彩子、葛西宏さん(前・産業医科大学教授)、田矢洋一さん(故人、前・国立がんセンター研究所部長)。
ビクトリア湖で種分化に挑む
SINEを発見したとき本当に知りたかったのはSINEの機能でした。しかし、2000年代の前半は、やっとヒトゲノムなどのデータが出てきたところで、分析技術も機が熟してはいなかったので、まず種分化を見ようと考えました。そこで短期間で種分化が起ったことで知られるシクリッドで研究を始めました。
シクリッドはスズキ目のカワスズメ科の淡水魚の総称です。脊椎動物の最大グループである魚類の約1割を占め、さらにその約7割がアフリカの大地溝帯の三大湖、タンガニーカ湖、マラウイ湖、ビクトリア湖にいて、それぞれの湖で別々に進化したと考えられています。ビクトリア湖は1万5千年前に一度干上がったとされ、短期間で一斉に種分化したのですから、種形成の原因探求には最適です。まずSINE法での系統解析を試みましたが、ビクトリア湖のシクリッドは近縁種でSINEの有無には差が見られず、ゲノムはほぼ同一とわかりました。それでも生態や色や形態が異なっているのですから、その差は自然選択された遺伝子の違いにあるはずです。ビクトリア湖は水深が浅い濁った不栄養湖ですから、現地の環境を見なくてはなりません。当初タンザニアには全くつてがなかったのですが、タンガニーカ湖で生態研究をしていた佐藤哲さん(現・総合地球環境研究所教授)を仲間に引き入れ、2004年の夏にタンザニアの水産学研究所(TAFIRI)との共同研究にこぎつけました。生態学者の溝入真治さんや形態学者の相原光人さんとチームを組んでの共同研究が始まりました。ビクトリア湖湖畔のムワンザに調査隊を派遣して、現地調査とサンプリングを行い、二万個体近いシクリッドを蒐集しました。標本にすると色が抜けるので、生体のカラー写真を撮影し、DNA抽出のための組織も採取しました。生態学と分類学の専門家との共同作業で、生態の記載と種の同定が厳密になされていますから、生きたサンプルを持ち帰ったようなものです。これを用いてさまざまなアプローチでシクリッドの進化研究を進めました。
ビクトリア湖のシクリッドの遺伝的背景は均一と考え、種特異的な変異が種の特徴を表す遺伝子であることに注目し、寺井洋平君(当時・東京工業大学助教)が遺伝子を探しました。そして赤から黄色の長波長側の光を吸収するオプシンであるLWS遺伝子に種特異的な多様性を見つけたのです。LWSの二つの対立遺伝子座対立遺伝子座父母に由来する2組の染色体(相同染色体)の同じ遺伝子座。がつくるタンパク質の光の吸収波長を調べ、濁った水域でやや深いところに棲む種では、赤系の長波長の光をよりよく受容できるようにシフトしていて、やや浅いところの種は青の方向にシフトしていることがわかりました。さらにオスの婚姻色を調べると、それと対応するような婚姻色を示しました(図13A)。行動実験からも浅瀬のメスは、淡青色のオスを選び、やや深場のメスは赤色のオスを選ぶことが確認できました(図13B)。これは感覚器適応種分化のモデルに合致します。光を受け取る感覚器が環境に適応して分化し、その分化した感覚器でよく見える婚姻色が次第に選択されることで交配相手が限定され、交雑が抑制されて性的隔離による種分化に至るのです。生息場所が近くでも、色覚の違いでよく見える相手が異なりますので、生殖隔離は維持されます。これは色と生態を記載したデータとDNAがあっての成果ですから、このプロジェクトだからこそできた研究だと喜んでいます。この研究はNatureのArticleとして出版され表紙にもなりました。ビクトリア湖のシクリッドに関しては、ほぼ全体をカバーするライブラリーができていますので、いつかビクトリア湖のシクリッド全体の進化を理解できると嬉しいですね。


ビクトリア湖の島でシクリッドのサンプル採集。手間はかかるが釣りが魚体を傷めなくて一番良い。右は生態学者の溝入真治博士。

ビクトリア湖をボートで沖にある島へ。後ろの女性は東工大の事務官。

図13A:水の透明度による種分化。より深場の濁った水では長波長(赤)がよく見えるタイプが分化し、婚姻色も赤になる。
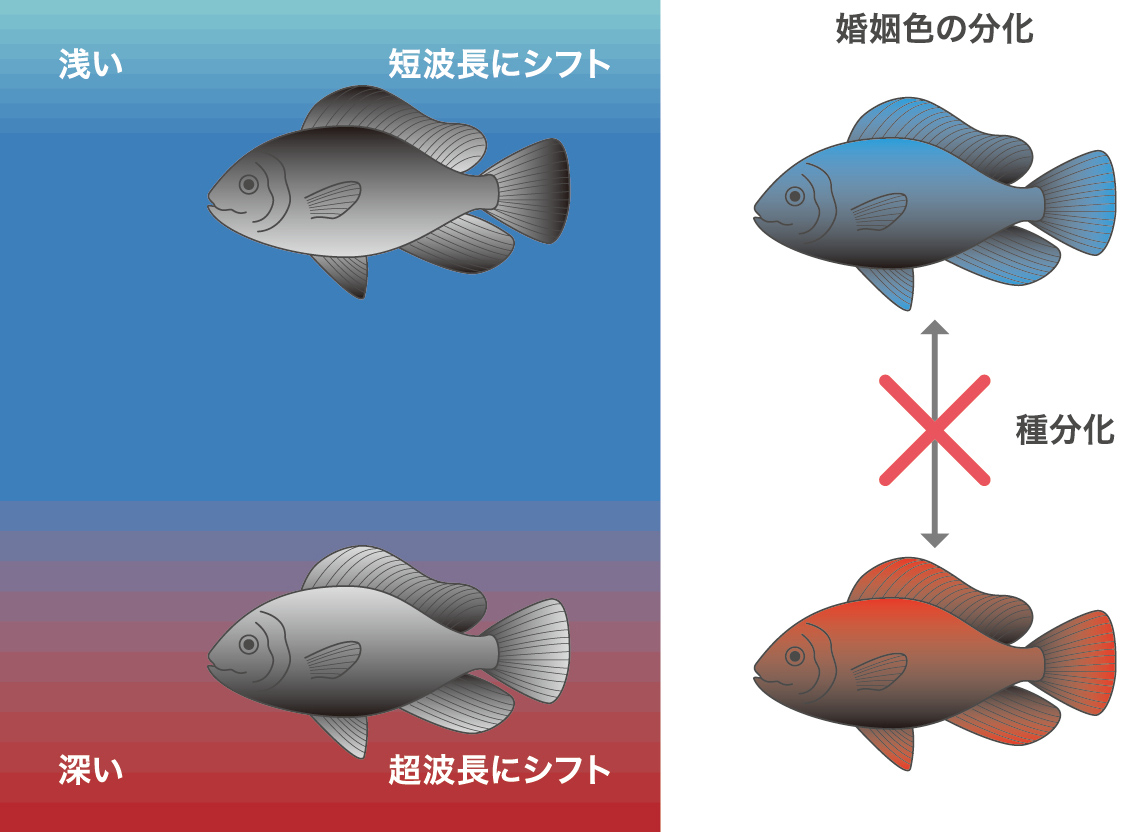
図13B:水深による種分化。浅瀬では青、深いところは赤に視覚も婚姻色もシフトする。

タンザニアからのお客様、右手前:キャサリーン・ムワコシャさん(タンザニア水産学研究所研究員)、右奥:ベン・ヌガツンガ さん(前・タンザニア水産学研究所総所長)、左手前: 相原光人(当時・国際科学振興財団研究員)、左奥:二階堂雅人(当時・東京工業大学助教)
シーラカンスがやってきた
タンザニアとの共同研究から思わぬ展開があったのがシーラカンスです。TAFIRIに共同研究の調印に行ったとき、ブワソンジ所長が、「タンザニアでシーラカンスが採れるんだが、興味があるか」と聞くので、「もちろん、是非そちらも共同研究させていただきたい」と答えました。その1ヶ月後に「シーラカンスを一頭寄贈するので、もう一度タンザニアに来る気はないか」とメールが届いたのです。サンプルがちょっともらえればいいくらいに考えていたので、半信半疑のまま、12月にタンザニアを再訪し、TAFIRIでの大統領臨席のパーティに臨みました。スワヒリ語なのでわかりませんが、大統領がシーラカンスを1匹日本に寄贈すると話したそうです。日本大使の同席の下、大臣にも確認してもらいました。ワシントン条約の締結や輸送手段の確保など複雑な手続きを経て、実際に冷凍のシーラカンスが東工大に到着したのは約1年後の2005年の11月11日でした。形態、機能、集団の視点から進化の学術研究を目的とする協定を結び、ワシントン条約発効後に初めて日本に輸入されたシーラカンスです。TAFIRIはヨーロッパの国とも共同研究をしていますが、日本人には親近感があるのでしょうか、シーラカンスをもらったのは東工大の岡田研究室だけです。その後、これを含めてシーラカンスの成魚5頭、稚魚10頭、卵6個が寄贈されました。そのうち2頭は、形態分析のためにCTを撮影し、解剖しました。卵を持つ個体の解剖は、魚類の専門家でいらっしゃる天皇陛下をお招きしました。この個体はシーラカンスの特徴である中空の脊柱や鰭にある骨格構造などが観察できるようにプラスチネーションを作成して、天皇陛下がご覧になったのと同じ状態で残してあります。残念ながら卵は全て未受精卵で発生過程を捉えることはできませんでしたが、今はゲノムが解っているので、実験で発生における遺伝子の機能を調べることもできます。

シーラカンスは発見者のスミスが、コモロ諸島を繁殖地としたので、その後、西インド洋の沿岸の広い地域で発見されていましたが、それが別の集団なのかコモロ諸島からの迷子なのかと意見が分かれていました。そこでまずシーラカンスの集団遺伝学をやりました。タンザニアとコモロのシーラカンスのミトコンドリアゲノムを解読し、先行研究でミトコンドリア解析が行われていた西インド洋の個体をあわせて72個体のミトコンドリアゲノムを比較しました。塩基置換のパターンから10個のハプロタイプハプロタイプ対立遺伝子座のどちらかの親に由来する組み合わせ。に分かれました。その出現頻度の分析から、コモロ諸島に特有なグループとそれ以外の2つに分かれる傾向が見られたのです。タンザニア北部とコモロ諸島の集団は少なくとも数十万年前には遺伝的交流がなかったという結果です。またコモロ諸島の個体ではハプロタイプの種類が少ないことから、おそらくワシントン条約発効以前にコモロ諸島の繁殖地からシーラカンスが乱獲されて、集団サイズが小さくなっていたのだろうと推測されました。タンザニアのシーラカンスの多様性を守ることがシーラカンスの保全に重要であるという我々の結果を受けて、タンザニア政府が沿岸にシーラカンスマリンパークを設置し、保護区を指定しました。これが保全に繋がれば、国際協力の成果です。タンザニアとの共同研究では、日本に来て一緒に研究したムジガニさんが岡田研究室で博士号をとりました。今はTAFIRIの所長になっています。科学技術外交にも貢献できたのではないでしょうか。
寄贈されたシーラカンスは進化を語る世界的にも貴重な試料ですので、共同研究を公募し、ゲノム解読には文部科学省の新領域「ゲノム支援」のサポートを受けてオールジャパンの体制で研究を進めました。個体を持っていますので、ゲノムを読むだけではなく、そこをスタートにいろいろな研究ができるはずです。損傷の少ない稚魚から冷凍のまま組織を取り分け、解析しました。ゲノムの専門家が協力して約27億塩基のゲノム配列が決まり、遺伝子数が2万2千から4千くらいとヒトに近い値がでました。
シーラカンスのゲノムは進化が遅いと言われていましたので、二階堂君が脊椎動物間で保存されている5千個以上の遺伝子のアミノ酸配列を使って系統樹をつくりました。するとやはりシーラカンスの枝が他の脊椎動物に比べて極端に短いことがわかりました。これは、アミノ酸置換速度が遅いことを意味します。次にインドネシアのシーラカンスとの比較で、塩基配列の置換速度を調べました。その結果、塩基置換のレベルでも遅く、ヒトとチンパンジーに比べて40倍も遅い のです。シーラカンスに分岐してから塩基置換速度が遅くなったため形態変化に関わる変異が起こり難くなり、化石種とほとんど同じ形をしているのではないかと推測しています。

2007年12月22日、秋篠宮さまをお呼びして、タンザニアからの1頭目のシーラカンスの解剖を行った。執刀は山田格さん(当時・国立科学博物館主任研究官、前列中央)。後列、左から五條堀孝さん(当時・遺伝学研究所教授)、秋篠宮文仁親王、一人おいて長谷川政美さん(前・統計数理研究所教授)、本人。
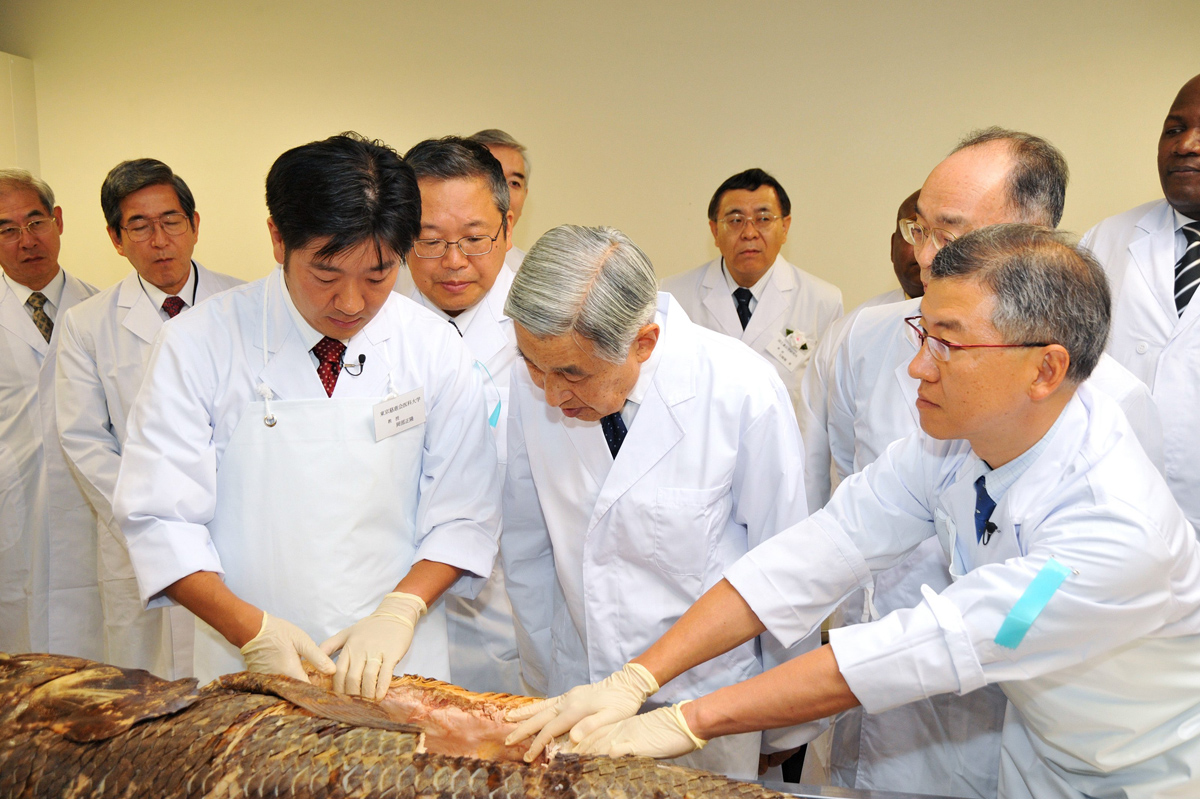
2009年10月5日、魚類がご専門の天皇陛下をお招きして、シーラカンスの解剖。執刀は岡部正隆さん(東京慈恵会医科大学教授:前列左)。その後ろが本人。

2011年の1月、NHKの人気科学番組、サイエンスZEROのロケを行った。番組出演者の安めぐみさんと研究室のメンバー。(後列左から)林由訓、吉田恒太、山口勝己、(安めぐみ)宮城竜太郎、岡田典弘(前列左から)二階堂雅人、工藤優、鈴木彦有、竹田正幸。

展示室には3匹の成体と1匹の稚魚と卵のシーラカンス標本があり、パネルで到着からこれまでの経緯や研究情報を解説している。
SINEが哺乳類の脳をつくった
シーラカンスのゲノムからSINEを探す過程で一つの大きな転機がありました。西原君がシーラカンスから新しいSINEを見つけたのです。このSINEファミリーと類似のファミリーがあり、それが哺乳類と爬虫類に分布しているので、有羊膜類(Amniotes)のSINE、 AmnSINE1と名付けました。AmnSINE1を哺乳類のゲノムで比べるとオーソログに相当する配列が哺乳類の非コード保存領域(CNE:Conserved Non-Coding Region)非コード保存領域遺伝子をコードしていないが、複数の生物種の間で配列が保存されているDNA領域。遺伝子発現の調節など遺伝子のはたらきに重要な機能をもつ配列を含む可能性がある。に存在し、哺乳類のゲノム中で機能を持ち、大進化に関わった可能性が考えられました。SINEの研究を始めたときから、SINEの存在する意味を考えてきましたが、いよいよそのときが来たと直感しました。哺乳類の共通祖先で増幅した配列が、ゲノムに分散し、それらが別々にほぼ同時に機能をもったということで、エグザプテーションバースト(外適応爆発)エグザプテーションバーストエグザプテーションは、外適応ともいい、ある形質や機能が、進化の過程で本来とは異なる別な機能を獲得すること。レトロポゾンについて使われる場合は、機能を持たない配列が新たに機能を獲得する現象をいう。エグザプテーションバーストはエグザプテーションが爆発的に一気に起きる現象。が起きているのではないかと想像し興奮しました。レトロポゾンが機能を持って大進化に関わる可能性が初めて示されたのです。そこで、ポスドクの佐々木剛君(現・東京農業大学教授)がマウス胚をつかったエンハンサーアッセイに挑戦しました。AmnSINE1が発生過程で遺伝子調節に関わるエンハンサー機能を持つかどうかの実験です。苦労の連続でしたが、1年以上の試行錯誤の末に明瞭に脳の特定の場所に活性のあるマウスがとれました(図14)。このデータを見たときの感動は忘れられません。このAS071と名付けた配列は、脳や四肢など発生のさまざまな過程で重要な役割をもつFgf8の近傍にあり、哺乳類の間で非常によく保存されていたのです。Fgf8遺伝子のプローブで10.5日の胚で発現を調べると、AS071がエンハンサー活性を示した間脳領域の位置とぴたりと一致し、しかも同じ位置にはニワトリでは発現が見られないという結果がでました。AS071座位のAmnSINE1がFgf8のエンハンサーであり哺乳類の脳の形成に関わる重要な機能を持つことは間違いありません。間脳領域のFgf8の機能はあまり知られていなかったのですが、理研の下郡朋美さんがFgf8が間脳にある視床での神経支配に関わり、髭の感覚を脳に投射するバレル構造の形成に影響するという論文を書いていることを知りました。早速相談すると、興味を持ってくれて共同研究が始まりました。下郡さんと共同研究できたことで脳に対する理解が格段に深まったと思います。哺乳類の視床に発現するFgf8が脳の機能マップの再構成に関わった可能性を示すことができました。他の領域でもエンハンサーアッセイを続け、他にも有望な活性がいくつか見つかりました。例えば、AS021です。この座位は13.5日の胚の大脳皮質に発現するSatb2のエンハンサーであることが証明されました。より詳細に細胞レベルでの発現を特定するため、そこで、今度は下郡さんに紹介してもらって、パリのジャック・モノー研究所のピエラニ博士に共同研究を申し込み、AS021がSatb2の発現制御を通じて、どのような哺乳類の高度な脳機能の獲得に関わるのか調べました。パリに滞在して実験に参加し、大脳の右半球と左半球を結びつける脳梁の特に腹側の形成に関わることを突き止めました。脳梁は哺乳類の脳にだけあり、除去しても生存に影響しないことから高度な脳処理に関わると考えられます。Satb2が脳梁形成に関わることは知られていたのですが、エンハンサー活性からニューロンまで特定したのは初めてです。AmnSINE1は哺乳類の共通祖先で増幅しそれが機能を持つことで保存され、哺乳類の進化に関わったことが証明されました。その後レトロポゾン由来のCNEの機能探索の論文はたくさんでてきましたが、発現だけでなく、機能まで調べた例はありませんでした。他にも哺乳類特有の二次口蓋の形成に関わる領域も見つかっています。哺乳類の進化は、ペルム紀末のP-T境界の大絶滅で陸上動物の3分の2以上が姿を消した時、急激に環境が変化する危機的な状況で生き残ったことがきっかけとされます。脳の発達や呼吸に有利な二次口蓋や横隔膜の獲得は生存に有利だったはずです。SINEによる新しいエンハンサーの獲得とそれによる哺乳類進化を地球の地質学的な環境変化(酸素濃度の低下など)と結びつけたのも、我々のオリジナルで、進化研究の幅を広げる役割を担ったと思っています。
CNEにあるAmnSINE1配列はまだ少なくとも百カ所以上あるのですが、この精度で全部の機能を個々に追求するのは不可能だと思ったところで定年を迎えました。哺乳類を生み出したネットワークが浮かび上がると面白いと思いながら、今は、国際科学振興財団ではゲノム全体を俯瞰してSINEの進化と機能のストーリーを作れないか考えているところです。反復配列はジャンクDNAといわれてきましたし、進化研究はお金にもなりません。基礎科学で研究者として生きていくには、自分の特技を生かし、最も重要なことにチャレンジすることが必要です。学問的に最も面白いと思うことをやり続けなければならないということです。30代後半で自分が発見したSINEに関し、疑問に思ったいくつかの問題に30年かけて一応自分の中では納得する答えを出せました。研究者として幸せだと思います。でもまだまだ終わっていません。僕は科学にエキサイトしていないと生きている気がしないんです。日本と台湾でこれからも続けますよ。

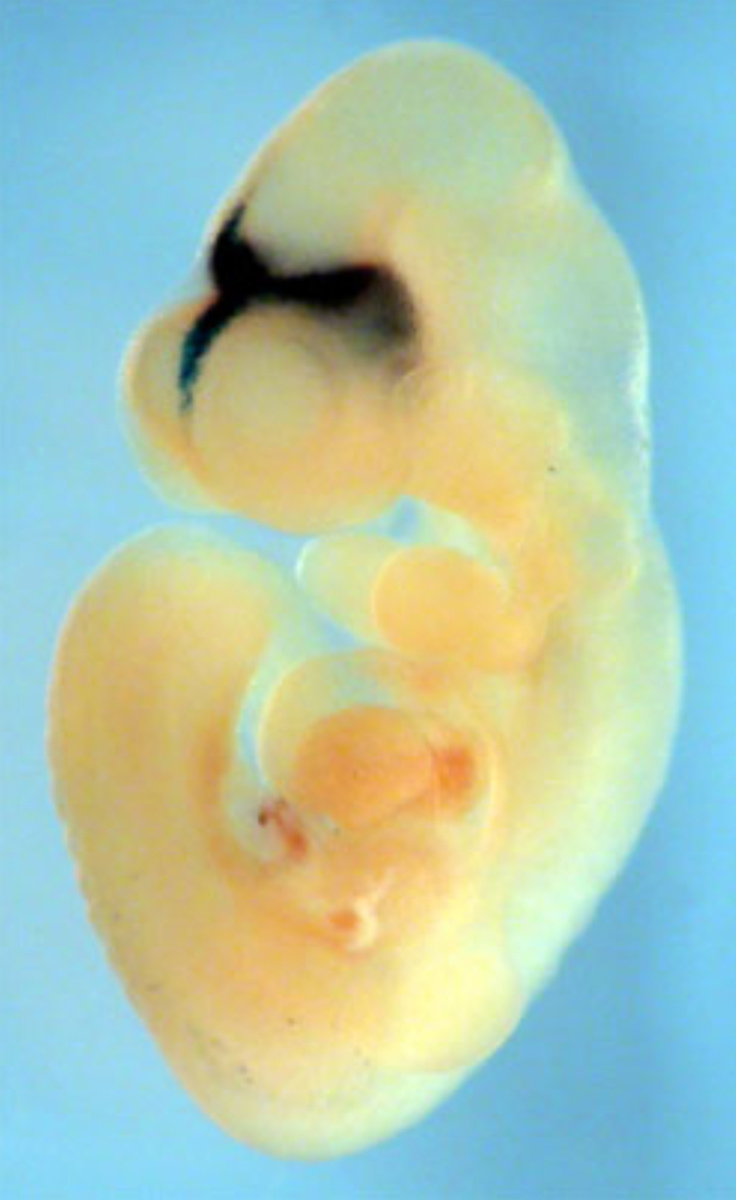
図14:AmnSINE1のエンハンサーアッセイ。10.5日のマウスの胚で発生中の間脳にくっきりと活性が見え、Fgf8の発現に一致した。

ピアニストの娘(右手前)のコンサートの後で会食。恩師の西村暹 先生(左奥)と開成の学友の山本正幸さん(左手前:当時・東大教授)が来て下さった。

65歳で東工大を定年になり、その後は日本ではつくばの国際科学振興財団の主席研究員(シーラカンス研究所所長)、台湾では国立成功大学の研究教授として活動中。国立成功大学(台湾)の生命研究科で。左から黄教授、蒋教授、本人。

2015年の夏に成功大学の学生さん達が日本神経科学会の年会で発表後、自宅でのパーティのスナップ。左から妻彩子、Sunny Sun, 本人、Chao-Ming Wang、Chiuan-Shiou Chiou、Chao-Li Huang、Chia-Wei Huang、Yang-Jung Yu、Yu-Ting Chiang、Yi-Wen Lin、Shih-Yu Lu、Simon Chu

娘夫婦と家族の近影。トイプードルのアルト君はもう15歳。




