



流行ではなく面白いことを
いろいろな生きものを見ていると、面白い現象に気づいてはっとする機会はいっぱい転がっています。博物学の時代に戻れというわけではありませんが、多様な生きものの生き様を見て、面白いテーマを考えることが今も大事だと思っています。
私の場合は、今の仕事の始まりは、飢餓状態に陥った酵母細胞を顕微鏡で見て、液胞の中で激しく運動している小さい粒を見たことです。当時誰も注目していなかった液胞に、酵母が飢餓を乗り切るための工夫があるのではないかと直感しました。これが実は、細胞のオートファジー(自食作用)を肉眼で確認した世界で初めての瞬間だったのです。顕微鏡をのぞくのが好きで、よく酵母を見続けていたからこそ、直感がはたらいたのだと思います。以来私は酵母を実験材料にして、オートファジーの仕組みをずっと追い続けてきました。
このような研究史を話すと、若い研究者が私の研究スタイルに憧れると言ってくれます。今は大学が疲弊して、研究も競争社会。若い人がこれから自分の研究を始めようという時、流行のテーマに乗っていないと不安に思わざるを得ない状況ですから。幸い私は競争とは無縁の世界で、他人に理解されなくともこつこつ研究を続けてこられましたが、今のような状況だったらきっと途中で止めざるを得なかったかもしれません。一人一人がよく考えてテーマを探し、多様なテーマを研究する事ができ、面白い新たな発見が出て来たらそれをきちんと伸ばせる研究環境を作らなければならないと痛切に感じています。

光学顕微鏡でとらえた出芽酵母のオートファジー。母細胞のほとんどを占める巨大な液胞の中に、小さい粒が多数見られる。
父と兄の期待
私は1945年、第二次大戦の終戦の年の2月に生まれました。4人兄弟の末っ子です。父親は大学の工学部に勤めていましたが、家は福岡市の外れにあり、農家の子どもたちばかりの中に大学の先生の息子がいるという環境でした。でも皆と一緒に自然の中で遊ぶのは楽しいものでした。年の離れた一番上の兄は終戦の年に政府が広島につくった科学教育のエリート中学の学級に選抜され広島にいましたが、原爆投下の直前に疎開していて危うく難を逃れました。そんな戦争の体験から兄は、大学は史学科に進みました。
父親は、末の子くらいは理系に進んでくれたらと期待はしていたようですが、何が何でも研究者になれというわけではありませんでした。東京の大学に入った兄は、休みに帰省する度に小学生の私に一冊の本を贈ってくれました。今考えてもそれらは優れた本で、宇宙、生物、化学の子供向けの本でした。八杉龍一の『生きものの歴史』や、ファラデーの『ろうそくの科学』、三宅泰雄の『空気の発見』などは、さし絵までよく覚えています。貧乏学生だったのにずいぶん無理をしてくれたんだと思いますがこれらの本との出会いから、大きな影響を受けました。私は昆虫少年で生きものが好きでしたが、なぜか化学に惹かれていて、高校でも化学部に属していました。

終戦の年、福岡市に生まれた。

4人兄弟の末っ子。上の兄は12歳年上。

小学生の時。父は九州大学の工学部教授だった。
新しい学科でのびのびと
父がいる九州大の工学部には行く気にはならず、理学部の化学に進みたいと思って東京大の理科II類に入学したのですが、教養学部時代の授業があまり面白くなかったので、自分が何をやりたいのかわからなくなってしまいました。幸運だったのは、教養学部の先生が中心となった基礎科学科が新設されたことです。科学の全分野を4年間かけて学び、その後に専門を選ぶという方針が気に入りました。新しい境界領域を目指すという学科の理念に共鳴した学生が集まり、1期生は元国立天文台長の海部宣男さん(現名誉教授)を始めとして、「幻の」とつくくらいユニークな人たちを輩出しました。私は2期生で、日立製作所でDNAシーケンサーを作った神原秀記君(現日立製作所フェロー)や、RNA研究の草分けとなった渡辺公綱君(現産業技術総合研究所研究技術統括)達が同級生です。
当時の教養学部の生物学には、筋肉研究の丸山工作先生(元千葉大学学長。故人)や生化学の茅野春雄先生(北海道大学名誉教授)、植物生理学の八巻敏雄先生がおられ、新しい学科で新しいことをやろうという雰囲気があふれていました。中でも今堀和友先生(現三菱化学生命科学研究所名誉所長)は日本における分子生物学の草分けとして、タンパク質の研究をされており、タンパク質の生合成の仕組みを解くという研究の旗印を立てておられました。セントラルドグマが確立していく生物学の大きな進展の中で、新しい学問分野への挑戦がとても魅力的に思えて、大学院は今堀研究室に行ったのです。
大腸菌でタンパク質合成の開始がどう制御されているかという課題に取り組み、博士課程に入ってからは大腸菌のタンパク質合成を阻害するコリシン毒素を研究するため、京都大学の生物物理学教室に内地留学しました。ちなみにこの教室も新設されたばかりの組織で、最初の卒業研究生には、近藤寿人(現大阪大学教授)や岡田清孝(現基礎生物学研究所所長)、坂野仁(現東京大学教授)など現在活躍中の若手が揃っていました。そんな経験から、大学は時々改組した方がいいというのが私の持論です。
大学院での研究はどれも、チャレンジングなテーマで実験も楽しかったのですが、実力が伴っていなかったのでしょう。たいした結果も出せず、博士論文を書き終わることなく、基礎科学科から農学部に移られた今堀先生の研究室に戻りました。「あなたは修士と博士で何をしたのか」と聞かれるとちょっとつらいのですが、自分が本当に面白いと思える研究をしているのだから構わないとのんびりしていました。実は学生結婚をして子どもも生まれていて、ワイフは創立されたばかりの三菱化成(現三菱化学)生命科学研究所でちゃんと働いていたのですが、私は論文も書かずにぶらぶらしていて、今から考えると本当にいいかげんだったと思いますね。

高校では化学クラブに所属。(本人:中列左から3人目)

東京大学に入学。下宿でリラックス。
Experiment: 季刊 生命誌 12号
原始生命はいかにしてたんぱく質をつくったか?
渡辺公綱

Scientist Library:
季刊 生命誌 24号
『筋肉をめぐる闘いの40年』
丸山工作

Scientist Library:
季刊 生命誌 20号
『昆虫の生きている仕組みを追う』
茅野春雄

Scientist Library:
季刊 生命誌 20号
『タンパク質と私の研究遍歴』
今堀和友

学部の卒業式で今堀先生(中央)を囲んで。先生の右隣は同級生の渡辺公綱君(現産業技術総合研究所研究技術統括)、左隣は本人。
酵母との出会い
博士課程を過ぎて2年後になんとか学位を取った後、今堀先生に「職は無いですか」と聞いてみました。「うーん、なかなかないから、海外でも行ってきたらどうだ。これからは細胞生物学の時代だから、ロックフェラー研究所がいいぞ」と、抗体分子の構造解析でノーベル賞を受賞したエーデルマンエーデルマン
Gerald Maurice Edelman
(1929 -)
アメリカの生物学者。1972年ノーベル生理医学賞受賞。の研究室を紹介して下さいました。そこでのテーマが、免疫細胞を活性化すると細胞分裂が誘導される仕組みを調べることと決まり、細胞周期に興味を持ち始めていたところだったので思いきって留学することにしました。
ところが行ってみると、エーデルマンは、免疫学をやめて発生生物学を始めると宣言していたのです。たぶん、別の分野でもう一度ノーベル賞を狙おうと考えたのでしょう。私のテーマも、マウス卵を用いた試験管内受精系の確立に変わってしまいました。技術自体は難しくはなかったのですが、その先どう展開したらいいのか答えが見えずと将来はないと本当につらい時期でしたね。その頃、カリフォルニア大学デービス校のハートウェルハートウェル
Leland H. Hartwell
(1939 -)アメリカの生物学者。2001年ノーベル生理医学賞受賞。が出芽酵母で細胞周期の変異体を分離し、分裂を制御する遺伝子を網羅的に解析するという研究をしていました。エーデルマン研最後の1年は、随分思い切って、出芽酵母を使って細胞増殖の制御機構をテーマにDNA複製の研究を始めることになりました。大腸菌から出発して高等動物の実験系に移って、自分は個体を見るよりも細胞の活動を分子レベルで扱う方が向いているなと改めて思いましたね。この酵母との出会いが、私の人生に重要な意味を持ってくるのです。

博士課程1年の時、「生化学若手の会」の夏の学校に参加(本人:中央)。右端の女性と1年後に学生結婚。

人のやらないことをやる
日本に帰る場所はないと思っていたのが、思いがけず東京大学理学部の植物学教室にいらした安楽泰宏先生(現名誉教授)から帰って来ませんかという話をいただきました。安楽先生とは、学生の時に生化学会の関東支部会で質問をしていただいた程度の面識でしたが、研究室のスタッフに外部の人間を積極的に採用する先生の方針から、今堀先生から私の現状を聞いて声をかけて下さったのです。
安楽先生は、大腸菌が細胞膜を介して物質を取りこむ膜輸送を研究し、世界的な仕事をされていました。しかし理学部の植物学教室では研究対象を大腸菌だけではなく、真核生物での研究を始めなければならないという意識をお持ちだったようです。「私は一生大腸菌をやるから、君は酵母を研究したらいい」と言われました。せっかく始めた酵母の研究ですから、それを続けられるのは嬉しかったのですが、研究室のテーマとあまりかけ離れたことをやるのもいいことではないという思いがありました。そこで沢山の方が手がけている細胞膜の輸送ではなく、細胞内の膜系で、まだ誰も注目していなかった液胞の膜輸送を始めることにしました。
これには実は伏線があります。エーデルマン研で酵母細胞から核を単離する実験をしていた時、捨てるつもりの上澄み液にきれいに濃縮されているオルガネラ(細胞小器官)の層があることに気づき、顕微鏡で見てそれが液胞だとわかり驚いたことがあったのです。私はもともと顕微鏡観察が好きで、酵母では光学顕微鏡で見ることのできる唯一のオルガネラである液胞を面白そうだと思っていました。
当時は植物でも液胞は不活性なオルガネラで細胞の中にあるごみ溜め程度にしか思われていませんでした。この液胞膜の物質輸送を研究テーマにすることを安楽先生に賛同して頂き自由に研究をさせてくれました。そして研究は意外な進展を見せ、液胞膜にはアミノ酸を始めとする能動輸送系と、プロトンを駆動するポンプとしてのV-ATPaseがあることを世界で初めて発見しました。液胞は不活性なオルガネラではなく、活発な膜輸送を介して細胞質の恒常性に積極的に関わっていることがわかったのです。これが、人のやらないことをやり、競争をしないで独自のものを出すという私のサイエンスのスタイルになりました。

安楽先生(後列右から2人目)の研究室の仲間と。米国留学中からヒゲを伸ばし始めた。(本人:前列右端)
「面白いことを見つけた!」
その後安楽研でも多くの人が酵母を研究するようになっていました。楽しい研究生活を10年あまり過ごしたのですが、そろそろ独立しなければと思いました。幸い教養学部で助教授のポストにつくことができました。その時、安楽先生の下でしていた仕事ではなく、新しいテーマを立てようと決心しました。スタッフが独立する時には学生さんを連れて行くという例もありますが、私の場合はまさしく単身での出発でした。当時持っていったものは、酵母の振とう培養器と滅菌装置、分光光度計と顕微鏡くらいでした。
この時に考えたテーマの1つが、液胞が細胞内の分解を担っていることを示し、その分子機構に迫れるかということです。液胞は細胞内のいろいろなものを貯め込んでいる一方、さまざまな分解酵素を含んでいる袋でもあります。液胞の内部で何をどのようにして分解されるのかは全くの謎でした。酵母の遺伝学は既に非常に進んでいましたが、私は遺伝学よりも顕微鏡をのぞいている方が性に合うという、酵母の研究者としては異端でしたから、なんとか液胞の中で起きていることを顕微鏡で観察できないかと考えました。
あるときひらめきました。酵母は栄養が無くなると、細胞内部をつくり変えて胞子を形成し、休眠状態で飢餓を乗り切ります。液胞が分解機能をもつとすれば、胞子形成時にその機能は最も活発にはたらいているにちがいありません。液胞内での分解を止めると分解される構造が見えるのではないかと考えました。さっそく酵母のストックセンターから液胞内の分解酵素が欠損している変異体を取り寄せ、飢餓状態で何が起きるか顕微鏡で観察するという非常に簡単な、しかし誰も思いつかなかった研究を始めました。研究の大事なアイデアは、ものがなく静かな環境で生まれるものだと思いますよ。
そして見つけたことは実に明確な現象でした。飢餓状態に3時間置くと液胞の中に小さな顆粒が段々蓄積し、激しく動き回っていました。細胞質の成分を取りこんだ膜構造ができて、それが液胞に取り込まれてブラウン運動をしていたのです。普段は動きがない酵母が見せる細胞内の動的な振る舞いに感動し、何時間も見つづけました。細胞の分解作用を初めて目で捉えたことが、オートファジーという細胞のリサイクルシステムを研究する出発点となりました。「とても面白いことを見つけた」と当時会う人ごとに話していたと、今になっても言われます。よほど嬉しかったのでしょう。
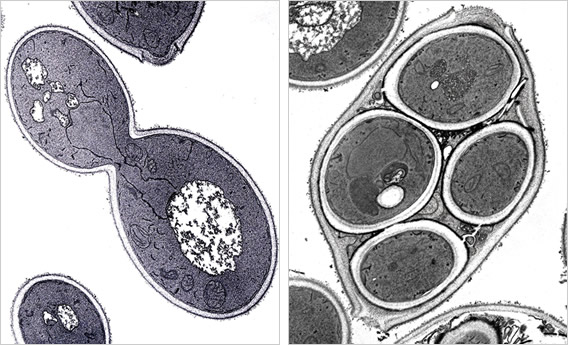
元気に生育中の酵母と、飢餓条件で誘導された胞子(減数分裂した4つの娘細胞)。
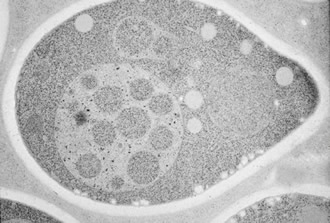
飢餓状態に置いたタンパク質分解酵素の変異体の電子顕微鏡写真。細胞内の大きな円が液胞で、中にあるのがオートファジックボディ。細胞質にはオートファゴソームが見られる。
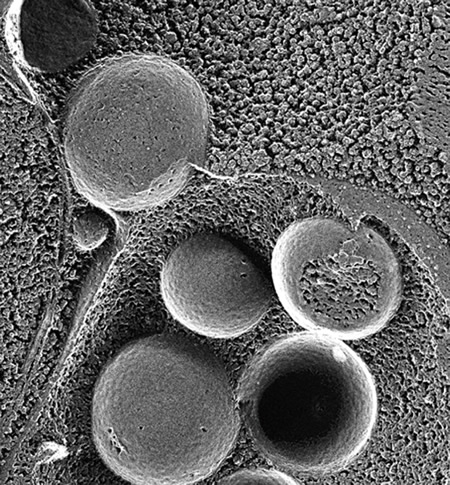
電子顕微鏡で捉えた、オートファゴソームが液胞に融合する瞬間のようす。

助教授として独立。両親が福岡から訪ねてくれた。
難産だった最初の論文
自分の見たものがオートファジーであることを証明するには、まず液胞に貯まる構造の実体をつきとめなければなりません。幸い、優れた電子顕微鏡技術を持つ馬場美鈴さん(日本女子大学)との共同研究でその過程を詳細に追うことができ、大きな成果が得られました。
飢餓状態の酵母では、分解すべき細胞質の成分を取り囲む二重膜構造が造られ、リボソームや時にはミトコンドリアまで丸ごと囲みます。これをオートファゴソームと呼びます。オートファゴソームは、外側の膜が液胞膜と融合し、細胞質を囲んだ内膜の球状の構造体が液胞の中に入ります。これが私が顕微鏡で見たブラウン運動する構造体で、オートファジックボディと名付けました。液胞内の分解酵素が正常にはたらく酵母ではこの構造体は瞬く間に分解されてしまうため、これまで誰も発見できなかったのです。
独立してから2ヶ月でオートファジーを見つけ、電子顕微鏡での観察を一通り終えたのが1年後です。そこでこれまでの成果を形態学的な記述と生理学的な過程の2つにわけて、別々の論文として投稿しました。ところが編集者が、1つの論文にまとめないと掲載できないと言うものですから、やりとりが長引き受理されるまで2年もかかってしまいました。その間にどういう経緯があったのか、なんと原稿のコピーがアメリカ中に広がり、何でこんな大事な現象を誰も見つけられなかったのかと騒ぎになったということをずっと後で人づてに聞きました。ただ、オートファジーの研究には電子顕微鏡が必須で我々の顕微鏡像がいかにも説得力があったので、すぐに分子生物学者が参入することはなかったのだろうと思います。
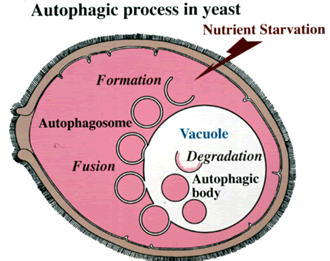
出芽酵母で起きるオートファジーの模式図。飢餓状態になると細胞内に二重膜構造が出現し、細胞質を囲むオートファゴソーム(Autophagosome)となる。オートファゴソームの外膜は液胞(Vacuole)膜と融合し、液胞に取りこまれた内膜はオートファジックボディ(Autophagic body)となって分解作用を受ける。
遺伝学から分子生物学へ
オートファジーに関する最初の論文が無事発表されましたが、その分子機構を明らかにするためにはどんな遺伝子が関わっているかを知る必要があります。そこでようやく本気になって遺伝学に取り組みました。最初に、飢餓状態でもオートファジックボディがたまらないという表現型を指標に、顕微鏡観察で変異体を見つけるという根気のいる方法でオートファジーが不能となるapg1変異株を単離することに成功しました。複雑なオートファジー現象に関わる遺伝子が1つだけとは考えられません。apg1変異体は、オートファジックボディを蓄積しないだけでなく、飢餓環境下での生存率が低くなることがわかりました。この表現型がオートファジーの欠損によると考えて、これを指標にしてもう一度変異体を選択しました。すると今度は、14種類のオートファジー変異株を得ることができました。この研究には大学院生の塚田美樹さんが大変大きな貢献をしてくれました。
現在わかっている主要なオートファジー遺伝子は全部で18個ですから、このスクリーニングが優れていて、後の展開を可能にするしっかりした基盤が出来た訳です。しかし、あとから参入してきた様々な酵母の研究グループがオートファジーに関わる遺伝子名それぞれをつけ始めたので、混乱を収拾するため命名を統一しようという動きになりました。私が優先権を強く主張すればよかったのですが、誰かの命名を生かすのではなく、新しくATGという名称をつけることに決まりました。しかし遺伝子番号に関しては APGが尊重され、ほとんどの遺伝子はAPGからそのままATGに読み替えれば良いということになりました。
こうしてオートファジーを発見し、ATG遺伝子を14個見つけることができたわけで、私個人として研究の最も楽しい時代だったかも知れません。

東京大教養学部の教官仲間。(本人:後の柱の前)
オートファジー研究の黄金時代
14個の遺伝子を手にして、次の課題はそれらの遺伝子にコードされるタンパク質の機能を明らかにすることです。東京大でも既にいくつかの遺伝子がクローニングできていましたが、本格的な研究の展開の契機は1988年に、スタッフ、研究設備などが充実した愛知県岡崎市の基礎生物学研究所に移ったことでした。助教授に吉森保さん(現大阪大学教授)、助手に野田健司さん(現大阪大学准教授)と鎌田芳彰さんを迎えました。
苦労すると思っていた遺伝子解析は、技術の発展に助けられました。遺伝子操作はどんどん簡単になり、酵母のゲノムプロジェクトも進んで14個の遺伝子も短時間で単離することができました。しかしそれらの機能に関しては、いずれも新規の遺伝子で、既知の遺伝子の構造から推定できることが全くない苦しい時期もありました。オートファジーには必須だが、欠損しても通常の細胞増殖に影響はなく、その機能は不明という報告が続くことになった訳です。
それぞれの遺伝子の機能は何か。機能解析の仕事に転機が訪れたのは、基礎生物学研究所に移って数年後のことでした。東京医科歯科大学で内科医をしていた水島昇さん(現東京医科歯科大学教授)がオートファジーの研究をしたいとやってきました。練習問題として、ちょうど帝京科学大学の大隅萬里子の研究室でクローニングしたばかりのAtg12の細胞内発現の解析をしてもらったのです。彼は短期間にAtg12がユビキチン化ユビキチン化オートファジーでは膜で囲った細胞質の中身がまるごと分解されるのに対し、エネルギー依存的にタンパク質を分解するユビキチン-プロテアソームシステムでは、プロテアソームという分解酵素の複合体がタンパク質を一つ一つ選んで分解する。この際、分解作用を受ける標的タンパク質にはユビキチンという小さなタンパク質が結合し、この過程をユビキチン化という。と同じ2段階の反応形式でAtg5と結合すること発見しました。Atg12が、当時注目され始めたユビキチン様タンパク質であることが明らかになった訳です。続いて、大学院生であった桐浴、一村両君が Atg8もまたユビキチン様タンパク質であり、驚くべきことに最終的にタンパク質ではなく膜リン脂質、フォスファチジルエタノールアミンに結合することを発見しました。オートファジーの膜動態に2つのユビキチン様の結合反応が必須であることは大きな驚きでした。
さらにその数年の間にAtg1キナーゼ、オートファジーに必須な PI3キナーゼ複合体などが相次いで見いだされ、Atgタンパク質の基本的な機能単位が明らかになりました。もう一つ特筆すべきことは、吉森、水島氏がATG遺伝子に関する哺乳類のホモログの研究を開始し、これらが酵母から人に至るまで保存されていることを示すことに成功したことが挙げられます。高等植物のATG遺伝子の同定も進み、現在私の研究室の出身者が、全国で動物や植物のオートファジー研究で活躍しています。
今から振り返るとAtg遺伝子の解析は、サイエンスには思いがけない展開があるという実例で面白い経験でしたね。新しい現象の発見とはまた違った体験であり、研究者人生としてはこの時がある意味で黄金時代だったと思います。

基礎生物学研究所に移り、オートファジー遺伝子の機能解析を進めた(本人:後列右から3番目)
Research: 季刊 生命誌 45号
オートファジーの現場をとらえる -細胞が自分を食べる理由-
水島 昇
オートファジー研究の現状
私たちが酵母でみつけたオートファジーのシステムは、すべての細胞に共通する現象です。オートファジーの研究は ATG遺伝子が見いだされたことで一気に高等動植物に広がり、ものすごい数の論文が発表される時代となって、病気との関連などが多くの関心を読んでいます。電子顕微鏡以外では詳細な観察が難しく、他の生物種での研究がなかなか進みませんでしたが、私達が作製したAtgタンパク質を認識する抗体や、 Atgタンパク質を蛍光標識する方法の導入など分子を見る技術が進みましたので、さまざまな細胞でオートファジーの過程を簡単にそして詳細に観察できるようになりました。
たとえばマウスでは、オートファジーが発生の初期や、新生児誕生、細菌感染防御に役割を演じていることが明らかになりましたし、脳でのオートファジーも調べられています。神経細胞は分裂しないので、細胞の中に古くなった異常タンパク質が貯まると薄まりようがなく、アルツハイマー病など神経変性疾患の原因となると考えられています。これをオートファジーが除去して防いでいるのです。
今やオートファジーは特殊な現象ではなく、うまくタンパク質をターンオーバーさせて細胞の質を保つという、細胞の持つ最も基本的な機能として理解できるようになってきました。一方でオートファジー研究が流行になって、細胞死、がん、老化など様々な高次機能にオートファジーが関わっているということをATG遺伝子のノックアウトで示し、オートファジーの新しい機能発見という論文になるといった、ちょっと乱暴な研究が目立ち始めました。それに比べて本格的にメカニズムを知ろうとする着実な研究は意外なほど少ないのです。
私自身は、オートファジーという現象の分子機構の解明に真正面から向き合うのが自分の歴史的使命と考えていて、それにはやはり酵母が一番適していると思っています。多くの知見が蓄積しており、色々な実験が可能で論理的に攻めることが可能だからです。私達は現在、オートファジーにおける膜動態、とりわけオートファゴソームという新規の膜構造がどのようにして造られるのかという問題、オートファジーの誘導機構、オートファジーによる分解の選択性など未だに解けていない根本問題をなんとしても解こうと励んでいます。オートファジーには多くの謎が隠されていて、それらを明らかにすることで、今まで知られていない細胞の大事な原理が明らかになるに違いないと思っていますし、現在議論されているさまざまな細胞内タンパク質分解の問題を理解する上で、むしろ近道だと考えるからです。
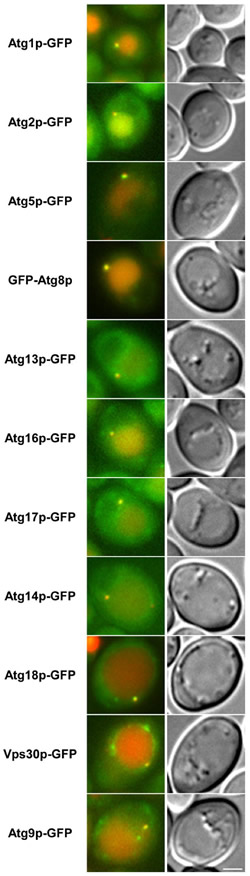
さまざまなAtgタンパク質に蛍光タンパク質GFPを結合し、細胞内での分布を調べた。
赤色は液胞の位置、緑色がAtgタンパク質の局在を示す。右の写真は光学顕微鏡観察での観察像。

オートファジーの最初の国際会議を岡崎で開催。

朝日賞の受賞式に夫婦で出席。
実験系にも多様さを
研究者にとって一番大切な環境は、当たり前ですが、落ち着いて研究を続けられるということです。ところが今の日本は非常に困った状況を抱えていて、それは科学行政の責任に違いありませんが、研究者の意識にも責任があるように思えます。
例えば若い研究者の中で、すぐに人に役に立つものか否かという考え方が近頃強くなっており、酵母よりマウス、マウスよりヒトの研究が重要だという意識が非常に強くなってきています。しかしガンを克服するのにガンの研究だけを進めればいいというわけではありません。生命の本質に迫る基本的で重要な問題は、大腸菌や酵母で解くことも大切なことです。さらに今や大学の研究予算の多くが成果主義で決まるという現実があります。マウスの研究・飼育には膨大な研究費がかかりますが、それをまかなえるのは一部の研究室だけでしょう。昔は大学が最低限保証していた講座費が、非効率な研究費のばらまきだとしてどんどん減らされていて、少ない予算でできる研究までも排除されています。サイエンスは効率だけを考えてよいものではありません。このままでは、研究の裾野はどんどん小さくなってしまいます。
社会に余裕がなくなり、それにつられてサイエンスも競争の世界になると本当の意味の科学する楽しさが失われてゆきます。若い人に考える余裕を与えず、早い者勝ちの世界で競争し続け、論文を量産しなければならない状態に追い込むことは科学を豊かにしてはくれません。研究者はじっくりとサイエンスと取り組もう、それにはお金がなくてもできるという開き直りや、一度や二度の失敗を恐れない覚悟が必要です。
大事な研究の始まりには必ずしも巨額な研究費は必要ではありません。しかし研究が発展、展開するためにはいつまでも研究費がなくて良い訳ではないことも自明の理です。日本の科学政策には、一度や二度の失敗も認め、いい仕事が現れたらきちんと拾い上げるシステムが必要なのです。意欲のある若い科学者を育てないと、日本の科学の未来は厳しいでしょう。

この春に研究室に入ったばかりの学生さんと。装置は近代化しても、酵母細胞をじっくり見るところから始めるのは変わらない。

この春に移った東京工業大学のメンバーで記念写真。








