年度別活動報告
年度別活動報告書:2002年度
脊索動物と節足動物の共通祖先を理解する 4-1 細胞間結合構造に基づいた動物の系統関係の推測
小田広樹(研究員)
秋山-小田康子(派遣研究員)
山崎一憲(奨励研究員)
入江雅美(派遣実験補助員)
はじめに
多細胞動物の系統関係は、100年以上もの間、多くの研究者によって議論がなされながら未だに結論の出ていない難問である1)。近年になって分子生物学的な手法がとられるようになり、解析方法に客観性が増してはいるものの、それらの方法にはまだまだ深刻な技術的な問題がある。
現在、最も広く行われている系統解析はDNAやRNAの塩基置換や蛋白質のアミノ酸置換に基づいたものであるが、この方法論が必ずしも万能でないことは多くの研究で指摘されている2)、3)。特に、動物の門や綱のレベルの(動物進化の初期で起こった)系統分岐を、この従来の方法で推測することは極めて危険である。それは、知りたい系統分岐が古ければ古いほど、置換速度のばらつきや絶滅によるデータの欠如等の影響を大きく受けてしまい、系統関係の確率論的な推測を困難にするからである。
最近は、このような従来の系統解析方法に代わって、長い進化の歴史で“稀”にしか起こらなかったであろう遺伝的な変化を見出すことによって、“深い”系統分岐をより確からしく推測する試みが行われつつある4)。例えば、ミトコンドリアゲノムでの遺伝子転移が昆虫類と甲殻類を系統的に結びつける証拠として示されており5)、その信頼性が認められている。このような「稀に起こった遺伝的変化」の探索は多細胞動物の門や綱レベルの系統関係を明らかにする上で必要不可欠であると思われる。
カドヘリン (正確には、クラシックカドヘリンとして定義される分子) は脊椎動物で最初に発見され、その後、ショウジョウバエや線虫、ウニ、ホヤからも単離された6)、7)。それぞれの動物でカドヘリン分子は形態形成に必須の細胞間接着分子であり、そのうちのあるメンバーは上皮細胞のアドヘレンスジャンクションと呼ばれる細胞間結合構造を担う中心分子である。私たちがカドヘリン分子に注目する最も大きな理由は、カドヘリンの分子構造が動物ごとに大きく異なっており、多細胞動物の進化過程でカドヘリンの構造が変化してきたことを示唆したからである8)。つまり、カドヘリンの構造変化が前述の「稀に起こった遺伝的変化」に当てはまる可能性があるからである。
そこで本研究は、その可能性を検証し、カドヘリンの構造的多様性が支持する多細胞動物の系統関係を提示することを目指した。その結果、カドヘリンの構造は小進化では確かに安定であるが、綱や門の境界で大きな構造的な違いが生じていることが明らかになり、カドヘリンの構造変化が「稀に起こった遺伝的な変化」に当てはまることが示唆された。そしてさらに、頭索動物が脊椎動物+尾索動物の姉妹群である可能性9) と、他の甲殻類に比べて鰓脚類が昆虫類に系統的に近い関係である可能性が示唆された。
結果と考察
(1)新たなカドヘリン遺伝子のクローニングとその構造決定
昨年度までに、昆虫類のフタホシコオロギ(Gryllus bimaculatus)とカイコガ(Bombyx mori)、甲殻類のヤマトヌマエビ(Caridina japonica)とフナムシ(Ligia exotica)、鋏角類のオオヒメグモ(Achaearanea tepidariorum)、軟体動物のケガキ(Sexostrea echinata)、棘皮動物のイトマキヒトデ(Asterina pectinifera)、半索動物のギボシムシ(Ptychodera flava)、頭索動物のナメクジウオ(Branchiostoma belcheri)からカドヘリン遺伝子をクローニングし、それらの構造を決定した。本年度はさらに、昆虫類のオオフォルソムトビムシ(Folsomia candida)、甲殻類のアルテミア(Artemia franciscana)からカドヘリン遺伝子をクローニングし、構造決定を行った。また、ナメクジウオからは新たに別のカドヘリン遺伝子をクローニングした。これらのカドヘリンの分子名と分子構造を図1にまとめた。
図1に示したドメインの中で、“EC”はカドヘリンリピートと呼ばれ、約110アミノ酸からなる繰り返し構造である。例外的に、ナメクジウオのカドヘリンはこのカドヘリンリピートを全く持たない。“CP”は細胞質領域で、すべてのカドヘリン分子に存在し、細胞骨格系との相互作用に重要な役割を果たしている。“NC”、“CE”、“LG”は異なる3種類のドメインで、その部分をまとめてPCCD(原始的なクラシックカドヘリンドメイン)複合体と呼んでいるが、この複合体は見つかっているすべての無脊索動物のカドヘリンに存在するが、脊椎動物と尾索動物のカドヘリンには存在しない。しかし、ナメクジウオのカドヘリンにはPCCD複合体の一部が存在する。
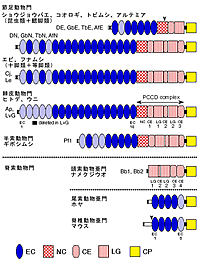 |
図1 カドヘリンの分子構造の多様性
図1に示したドメインの中で、“EC”はカドヘリンリピートと呼ばれ、約110アミノ酸からなる繰り返し構造である。例外的に、ナメクジウオのカドヘリンはこのカドヘリンリピートを全く持たない。“CP”は細胞質領域で、すべてのカドヘリン分子に存在し、細胞骨格系との相互作用に重要な役割を果たしている。“NC”、“CE”、“LG”は異なる3種類のドメインで、その部分をまとめてPCCD(原始的なクラシックカドヘリンドメイン)複合体と呼んでいるが、この複合体は見つかっているすべての無脊索動物のカドヘリンに存在するが、脊椎動物と尾索動物のカドヘリンには存在しない。しかし、ナメクジウオのカドヘリンにはPCCD複合体の一部が存在する。
今までに明らかになったカドヘリンを整理すると、昆虫類と鰓脚類のアルテミアではそれぞれ2つのタイプのカドヘリンが得られており、それらはショウジョウバエのDEカドヘリンとDNカドヘリンに対応した。しかしながら、アルテミアと同じ甲殻類に属しているヤマトヌマエビ(十脚類)とフナムシ(等脚類)や鋏角類のオオヒメグモではDNカドヘリンに対応する分子しか得られていない。ヒトデのApカドヘリンは、ウニのLvGカドヘリンとほぼ同じ構造をしており、これらはDNカドヘリンとも全体的構造が似ていた。棘皮動物のカドヘリンと比べた場合、半索動物ギボシムシのPf1カドヘリンは構造的に短縮化されていた。ナメクジウオに少なくとも2つのカドヘリンが存在したが、どちらもカドヘリンリピートを持たず、細胞外領域はPCCD複合体だけからなっていた。尾索動物(ホヤ)では、最近公表されたCiona intestinalisの全ゲノム情報を調べるかぎりでは、2つのカドヘリン遺伝子しか存在せず、構造はいずれも典型的な脊椎動物タイプであった(細胞外領域は5回のカドヘリンリピートのみからなる)。
これらの様々な動物のカドヘリンの分子構造が示唆する重要な点は、系統的に当然近いと思われる動物間では全体的構造が非常によく保持されているが、ある動物群とある動物群を境界にしてその構造が大きく異なっていることである。このことは、カドヘリン遺伝子の構造的多様性が「稀に起こった遺伝的変化」の結果である可能性を強く支持した。
(2)カドヘリンの発現パターン及びアドヘレンスジャンクションへの局在の解析
それぞれのカドヘリンが動物の形作りにどのように関わっているかを知るために、私たちはいくつかのカドヘリンに対する抗体を作製し、その蛋白質の発現を調べた。その結果、フタホシコオロギのGbEカドヘリン、フナムシのLeカドヘリン、ヤマトヌマエビのCjカドヘリン、ギボシムシのPf1カドヘリン、ナメクジウオのBb1カドヘリンが少なくとも胚の外胚葉性上皮で発現しており、アドヘレンスジャンクションに局在していることが明らかになった。ナメクジウオのBb2カドヘリンは中胚葉及び内胚葉で発現していた。これらの発現の結果と前項目で示した構造の結果を考え合わせると、上皮のアドヘレンスジャンクションを担うカドヘリンの分子構造が大進化の過程で様々に変化したと推測された。
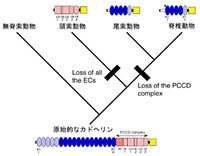 |
図2 推測されるカドヘリンの構造変化と脊椎動物の系統関係
(3)脊索動物の系統関係と昆虫類/甲殻類の系統関係
昨年度の報告書でも触れたが、様々な動物のカドヘリンの構造的な関係は、脊索動物の系統関係に関して、意味のある情報を提供する。一般的には、頭索動物が最も脊椎動物に近い脊索動物であると考えられている 10)、11) が、実は今までのところ、十分に強固な分子的な根拠は報告されていない。このような中で、私たちのデータは頭索動物ではなく尾索動物が脊椎動物に最も近い脊索動物であることを支持する9) 。推測されるカドヘリン分子の進化のプロセスを図2に示した。
左右相称動物の原始的なカドヘリンは、現在の棘皮動物や甲殻類で見つかっているような、細胞外領域に多数のカドヘリンリピート(たぶん18回)とPCCD複合体を持っていたと推測される。脊椎動物+尾索動物の系統ではPCCD複合体が完全に失われ、頭索動物の系統ではカドヘリンリピートが完全に失われたようだ。似たようなカドヘリンの構造的欠失は、半索動物の系統や昆虫類+鰓脚類の系統(鰓脚類の系統的な位置に関しては後述)でも独立的に起こったようである。しかし、このような私たちの解釈は、実際に進化の歴史上で起こった遺伝子重複や構造変化を極端に単純化している可能性があり、当然、まだまだ検討の余地がある。
最後に、前述の鰓脚類の系統的な位置に関してであるが、この動物群は一般的には原始的な甲殻類であるとされている。しかし、私たちは鰓脚類のアルテミアから、昆虫類でしか見つかっていない、DEカドヘリンに対応する遺伝子を単離した。このカドヘリンが昆虫類と同じように上皮のアドヘレンスジャンクションを担っているかは確認できていないが、他の甲殻類(十脚類と等脚類)ではDNカドヘリンに似た構造のカドヘリンが上皮のアドヘレンスジャンクションを担っていた。これらのデータは、鰓脚類が十脚類や等脚類よりも昆虫類に近い可能性を示唆した。
おわりに
私たちは多細胞動物の系統関係をカドヘリンの分子構造を指標にして調べ、現在広く受け入れられているものとは必ずしも一致しない系統関係を得た。私たちの斬新な系統関係がもし本当に正しいならば、この系統関係と矛盾しない進化的な変化をさらに見出すことができるはずである。現在すでにマウスやフグ、ホヤなどのゲノムプロジェクトが完了しているが、今後も脊索動物のゲノム情報はナメクジウオを含め様々な種で明らかになってくると予想される。おそらく、脊索動物の系統関係は近いうちに結論がでるはずである。それに対し、昆虫類と他の節足動物の系統関係はゲノムプロジェクトによる進展は短期的には期待できないであろう。今後の解析は節足動物の系統関係に焦点を絞って行っていきたい。
最後に、本研究の中でナメクジウオとギボシムシの解析は、和田洋博士(京都大、瀬戸臨海)、Shicui Zhang博士 (青島海洋大)、田川訓史博士(ハワイ大)、佐藤矩行博士(京都大)、Tom Humphreys博士(ハワイ大)との共同研究で行った。