年度別活動報告
年度別活動報告書:2001年度
脊索動物と節足動物の共通祖先を理解する (1) 細胞の構造と機能の解析 (2) 発生メカニズムの解析
小田広樹(研究員、代表者)
秋山-小田康子(派遣研究員)
はじめに
ヒトを含む脊索動物門と昆虫を含む節足動物門は、地球の様々な環境に適応した、学問上極めて重要な動物門である。この二つの動物群は5億年以上前に分岐し、長い歴史の中でそれぞれ独特な進化を遂げた。脊索動物と節足動物の進化の歴史は比較的豊富な化石記録によってぼんやりとは知ることができるが、「脊索動物と節足動物がどのような動物からどのように変化して誕生したのか?」という問題に関しては、化石は何も語ってくれない。私たちはこの問題を解く手がかりを得るために、分子生物学的手法を用いた解析を現存の動物に対して行っている。それは、動物系統によって異なった変化をしながら引き継がれてきた遺伝情報の中に、脊索動物と節足動物の起源を知る手がかりを探すことができるのではないかと考えているからである。
私たちは研究に2つの方向性を持たせている。第一は、動物の進化過程で起こった「細胞の構造や機能の変化」を調べることである。第二は、動物の進化過程で起こった「発生メカニズムの変化」を調べることである。この2つの観点から動物を比較することによって、どの動物群とどの動物群の間でその変化が起こったのかを明らかにしていくのである。つまり、比較細胞生物学と比較発生生物学を行うことにより、動物の歴史の解明を目指す。
この研究の足がかりとなるデータは、平成13年9月まで在籍していた科学技術振興事業団、月田細胞軸プロジェクトに於いて、小田広樹と秋山-小田康子が中心になって行なった研究より得たものである。本報告書ではまずこれを「背景」として簡単にまとめ、引き続いて、平成13年10月1日からBRHで行った活動について報告する。
背景
背景1-1 アドヘレンスジャンクションを担う分子カドヘリン
多細胞動物は、細胞と細胞がつながった細胞の集合体として体の形を作っている。細胞と細胞は特殊な結合構造 (ジャンクション) でつながっており、何種類かあるジャンクションの中でも、アドヘレンスジャンクション (図1) は最も体の形作りに重要なジャンクションである。アドヘレンスジャンクションで細胞と細胞の連結を担う分子がカドヘリンであり、この分子は隣り合った細胞間でホモフィリックな結合をする。
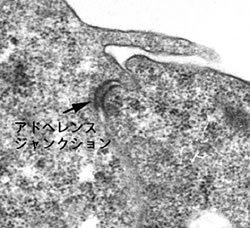 |
【図1】
背景1-2 カドヘリンの分子構造の多様性
カドヘリン (正確には、クラシックカドヘリンとして定義される分子) は脊椎動物で最初に発見され、その後、ショウジョウバエや線虫、ウニ、ホヤからも単離された。そこで分かってきたことは、カドヘリンの分子構造が動物ごとに異なっていることであった。このことは動物の進化過程でカドヘリンの構造がダイナミックに変化してきたことを意味する。しかしながら、進化の過程でどのような変化が起こったのかを推測するにはまだあまりに情報が乏しかった。そこで私達は、幅広い動物分類群からカドヘリン遺伝子をクローニングし、構造に関する情報を得た。実際に扱った動物は、昆虫類からコオロギとカイコ、甲殻類からエビとフナムシ、他の節足動物からクモ、軟体動物からケガキ、棘皮動物からヒトデ、半索動物からギボシムシ、頭索動物からナメクジウオである。結果を図2にまとめたが、脊索動物と無脊索動物の間でカドヘリンの構造に明確な違いがあった。
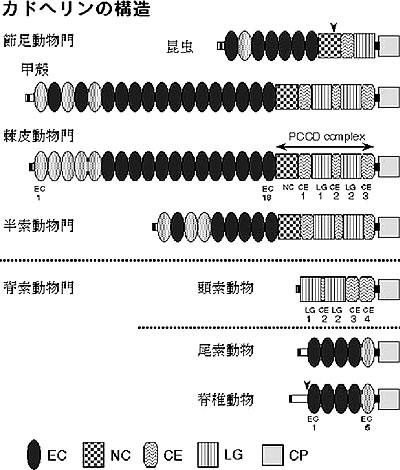 |
【図2】
背景1-3 ナメクジウオのカドヘリンが示唆する脊索動物の系統関係
最も興味深い発見は、脊索動物門の頭索動物ナメクジウオ (Branchiostoma belcheri) のカドヘリンが非常に奇妙な構造をしており、カドヘリンに特徴的な細胞外領域の繰り返し構造が全くなかったことである(上図)。私達はこの奇妙な構造をしたナメクジウオのカドヘリンが細胞間接着分子として機能できることを示し、抗体の作製によりアドヘレンスジャンクションに局在することも確認した。そしてさらに重要なのは、ナメクジウオのカドヘリンの細胞外領域が、他の脊索動物のカドヘリンとは全く似ていないが、無脊索動物のカドヘリンとは細胞外領域にも確かに類似性があったことである。このことは、尾索動物は最も原始的な脊索動物であり、頭索動物が脊椎動物に最も近い動物であるとする系統関係に関する常識とは矛盾した。つまり、頭索動物は尾索動物よりも原始的な脊索動物であるという可能性を示唆した。しかし、これはあくまでも可能性であり、それぞれの亜門で独立的に構造変化が起こった可能性も排除できないし、それまで複数種あったカドヘリンのうちいくらかをそれぞれの亜門で独立的に失った可能性も排除できない。現在いくつかの脊索動物で始まっているゲノムプロジェクトが進展することによって、様々な遺伝子や分子に関する解析が可能になることが予想される。今後、脊索動物の系統関係に関する議論も活発になってくるに違いない。
背景1-4 節足動物におけるカドヘリンの進化
昆虫綱双翅目のキイロショウジョウバエ (Drosophila melanogaster) では構造の異なる2種類のカドヘリンが存在し、ひとつは上皮で発現し(DEカドヘリン)、もうひとつは中胚葉や神経で発現する(DNカドヘリン)。私達は鱗翅目のカイコ (Bombyx mori) や直翅目のフタホシコオロギ (Gryllus bimaculatus) を調べたが、それぞれの昆虫からショウジョウバエで分かっているカドヘリンに対応する2種類のカドヘリンを見つけた。続いて、甲殻綱のフナムシ (Ligia exotica)とヤマトヌマエビ (Caridina japonica) からもカドヘリン遺伝子のクローニングを試みたが、昆虫の神経で発現するタイプのカドヘリンと構造的に類似したものをそれぞれの動物から1種類を得ただけであった。つまり、昆虫の上皮で発現するタイプに構造が類似したカドヘリンは甲殻類では見つからなかった。
さらに鋏角綱のオオヒメグモ (Achaearanea tepidariorum) からもカドヘリン遺伝子のクローニングを行ったが、得られたものは甲殻類のカドヘリンに構造が類似していた。昆虫の上皮で発現するタイプと類似するものは、クモからも得られなかった。
節足動物のカドヘリンに関しては、現段階では断定的なことは言えないが、昆虫の起源か、もしくはその前後で何らかの変化が起こった可能性がある。
背景2-1 クモの分子発生生物学の意義
この20年の間で、昆虫綱に属するショウジョウバエに関する遺伝学的、分子生物学的研究が飛躍的に進み、この動物の胚発生を支配する分子基盤がかなり詳しく明らかになった。しかし、ハエのことを知っただけで節足動物の祖先のことを理解できるかというと、全くそういうわけではない。節足動物の多様化の過程で、ハエが祖先的な形質をどれだけ失い、どれだけ特殊性を獲得したかを知らなければならない。そのために私達は、ハエから系統的に離れた節足動物である鋏角綱のオオヒメグモ(図3)を研究対象とし、この動物の胚発生を分子生物学的手法を用いて調べる。クモとハエの比較により、節足動物の祖先のことを少しでもよく理解できるようになることを期待している。
 |
【図3】
背景2-2 クモの胚発生に関わる遺伝子のクローニングと発現解析
ショウジョウバエの胚発生に関わる遺伝子と相同なクモの遺伝子をクローニングし、その遺伝子の発現パターンをwhole mount in situ hybridization法で調べた。上の写真にはその一部を示すが、様々なパターンでクモ胚が染め出された。これらの染色パターンは、クモとハエの胚発生を比較するための貴重なデータである。例えば、
engrailed (en) の発現パターンはクモとハエで本質的な違いはないと思われる。それに対しforkhead (fkh) は、ハエでは主に腸と卵黄で発現しているが、クモでは腸と卵黄に加えて正中線に沿った表層外胚葉にも発現しており、ハエとクモの違いが観察された。このような比較を進めていくことによって、共通点と相違点を明らかにすることができ、節足動物の祖先的な形質を理解するための手がかかりが得られると期待している。
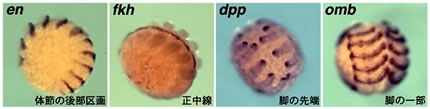 |
【図4】
結果と考察
研究室の立ち上げ
10月1日に着任してから約2週間かけて、研究室のセットアップを行った。実験機器等の多くは前所属先の科学技術振興事業団から借り受け、研究室もしくはオープンラボに搬入した。前任者から引き継いだ機器や器具類は出来る限りそのまま使用するようにした。必要な機器や試薬類を新たに購入して、11月から少しずつ実験ができるようになった。
甲殻類のカドヘリンの発現解析(多くは科学技術振興事業団で行った)
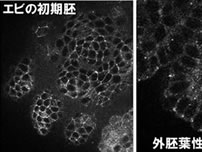
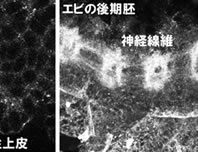
私たちがクローニングした甲殻類のカドヘリンは、中胚葉と神経に発現する昆虫のカドヘリンと構造的に類似していたわけであるが、甲殻の胚でも同じように発現しているのだろうか?このことを調べるために、フナムシのカドヘリン (Leカドヘリン) の細胞外領域にあるリピート構造に対して得た抗血清をアフィニティー精製したものを用いて、フナムシ胚を染色した(図5左の写真)。その結果、外胚葉性上皮の細胞境界でアドヘレンスジャンクションが存在すると考えられる部分がきれいに染色され、昆虫とは状況が異なることが示唆された。さらに、抗Leカドヘリン抗血清を用いてエビ胚を染色すると、同じように上皮細胞が染色され、それに加えて、後期胚では神経細胞の神経線維がきれいに染色された(図5右の写真)。これらの結果により、私たちがクローニングした甲殻のカドヘリンは、同じような構造をしている昆虫のカドヘリンとは異なって、ユビキタスな発現を示すものであることが分かった。つまり、昆虫と甲殻では外胚葉性上皮で発現しているカドヘリンが構造的に違うものであることが示唆された。この違いがどのようにして生じたのかを、さらに他の節足動物を調べることで明らかにする必要がある。もしかすると、昆虫の起源に迫ることができるかもしれないと期待している。
|
|||
| 【図5】 | |||
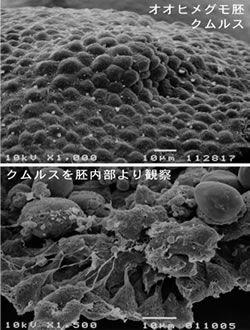
クモの体軸形成に関わると思われるクムルスの解析
クモとハエは卵の形から大きく異なる。クモの卵は外見上全く対称な球形であるのに対し、ハエの卵はかなり非対称な形をしており、外見だけからどちらが前で後ろか、どちらが腹で背中かを知ることができる。一方、クモ胚で前後、背腹の向きが外見でわかるようになるのは胚発生がいくらか進んでからである。その体軸が明らかになる最初の現象は、クムルスの出現と移動である(図5の写真)。私たちはこのクムルスを走査型電子顕微鏡で観察した。クムルスは細胞層の肥厚として観察されるが(図6上の写真)、地球に例えれば北極にあたる部分に形成され、体の後方に向かって約6-7時間かけて赤道付近にまで移動する。クムルスの隆起した細胞層の内側には、約10個の間充織様の細胞(仮にクムルス内部細胞と呼ぶ)が存在し、この細胞に向かって表上皮細胞の基底面から細長い突起が伸びていることがわかった(図6下の写真)。上皮細胞層の基底面を移動するクムルス内部細胞によって突起形成が誘起されているようであった。一方、クムルス内部細胞もアクチン繊維を含む突起を上皮細胞の細胞間隙を縫うように伸ばしていることがファロイジン染色で明らかになった。これらの観察から、クムルスの移動に際して、表層上皮細胞とクムルス内部細胞との間で緊密な情報のやりとりが行れていることが推測された。
 |
| 【図5】 |
私たちが科学技術振興事業団でクローニングしたクモのDpp相同遺伝子(AtDpp) は、クムルス内部細胞で特異的に発現していた。ショウジョウバエでは、Dppは背側を誘導する重要な分泌型シグナル分子であることが知られている。このことから、クモ胚ではAtDppを介した細胞間のコミュニケーションが表層上皮細胞とクムルス内部細胞の間で行われている可能性が考えられる。ここで最も興味深いことは、そのシグナル分子を発現している細胞の性質やふるまいがクモとハエで極端に異なっていることである。ハエではDppを発現する細胞は上皮形態を終始維持しており、胚内部に入って長い距離を移動するようなことはない。今後、この違いが意味するところをさらに詳しく調べる必要がある。
 |
| 【図6】 |
おわりに
以上のような研究を通して、私たちは動物の大進化のメカニズムを解明したいと考えている。大進化とは、動物の「門(節足動物門など)」や「綱(昆虫綱や甲殻綱など)」のレベルの進化を意味する。大進化の研究は、動物間であまりに違いが大きすぎるために論理的な比較検討が満足にできないという理由で、今まで敬遠されがちな研究分野であった。その一方で、5億年から6億年以上前に起こった動物界の変化を推測し、議論することは科学者にとって大きな魅力である。このような中で、近年の分子生物学の発展とそれに伴う遺伝情報の蓄積は、限られた動物種(ヒト、ショウジョウバエ、線虫など)に対するものではあるが、大進化の研究を急速に現実的なものとさせた。私達は、脊索動物と節足動物に注目しながら動物界の歴史の大きな流れを明らかにすることを目指すととともに、生物が営んできた「とてつもなく長い時間」をできるだけ多くの人々に感じてもらうための努力をしていきたいと考えている。