年度別活動報告
年度別活動報告書:1999年度
DNA から見たオサムシの多様化
蘇 智慧(研究員)
大澤省三(顧問)
金 衝坤(奨励研究員)
岡田節人(館長)
中村桂子(副館長)
はじめに
甲虫の仲間であるオサムシは, 主として北半球に分布し, ごく一部が南米 (チリオサムシ) とオーストラリア (オーストラリアオサムシ) に産する。世界に約1000種, 日本には35種がいる。この仲間のほとんどは, 後ろばねが退化していて飛べず, 歩いてしか移動ができない。そのため, 移動能力が制限され, 地理的変異が出やすく, 生物の多様化, 種分化, 地史と関連した分布圏の成立などを考察するのに格好の材料となる。これらの問題へのアプローチとして, われわれは主としてミトコンドリア(mt) DNA, また補助的に核 28S DNA と ITSを使って, 全世界のオサムシの系統関係を調べてきた。本研究プロジェクトの終了を迎える今年度はこれまで得られた結果を総合考察し紹介する。なお, 今回は過去のオサムシ研究の総括ともなるので, かなり長い報告とした。
材料
これまで集めた DNA分析用サンプルの採集地点を世界地図の上にブロットしてみたのが図1である。世界のおよそ35ヶ国から集めてきた標本数はすでに 2000 点を越え, 採集地点も 500地点を越えている。内容的にはオサムシ亜族を構成する 95% 以上の亜属, 半分以上の種類を入手しており, オサムシの全分布域をカバーしている。
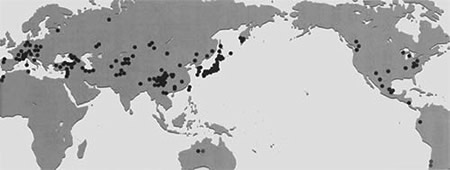 |
【図1:DNA分析用サンプルの採集地点】
ごく近い産地同士の場合、ひとつの黒丸が複数の地点(最高十数地点に及ぶ)を表す。
また、南米のチリオサムシ、豪州のオーストラリアオサムシは本図から省いている 。
結果と考察
mtND5 遺伝子の進化速度
分子系統樹からの分岐年代の推定は,化石や地史のデータを必要とするのでかなり難しい。だが,年代スケールの設定は系統進化研究の最も重要なポイントの1つで,これがないと系統的データの解釈が大幅に限定されてしまう。
日本特産のアキタクロナガオサムシの亜種ホソアオクロナガオサムシ (Euleptocarabus porrecticollis kansaiensis) は近畿全域に分布するが,淀川を挟んで東側のものと西側のものは mtND5 遺伝子の塩基配列で明らかに区別できる。この東西 2つの集団は, 約 300 万年前に完成した淀川水系による隔離で分岐したと思われる。もう1つ日本の特産種マイマイカブリ (Damaster blaptoides) の系統は日本列島の形成と関連している。古地磁気の研究から,古代日本は約1500万年前にユーラシア大陸の東縁部から分離し,東日本と西日本に分かれたとされている。マイマイカブリの祖先もこの時に東系統と西系統に分かれたと推定される。さらに,ヨーロッパに広く分布しているクロツヤオサムシ (Phricocarabus glabratus) やヒメダルマオサムシ (Tomocarabus convexus) はアルプスを挟んで北の集団と,南の集団が mtND5 遺伝子では明確に区別できるので,これらのオサムシの2系統はアルプスの隆起による地理的隔離によって形成されたと推定される。以上の地史のデータからオサムシのmtND5 遺伝子の進化距離 (D) 0.01 ユニットは約 360 万年に当たると推定された 。
オーストラリアオサムシとチリオサムシの系統的位置と起源
オサムシは分類学的にはオサムシ亜科 (Carabinae) に属し, セダカオサムシ族 (Cychrini) とオサムシ族 (Carabini) に分けられ, オサムシ族はさらにカタビロオサムシ亜族 (Calosomina) とオサムシ亜族 (Carabina) に分割されている 。mtND5 遺伝子による系統解析の結果は上記の高次分類を支持している。しかし, Carabinae に属するチリオサムシ (Ceroglossus) とオーストラリアオサムシ (Pamborus) の系統分類学的位置については定説がなく, 未知である。オーストラリアオサムシについては Kryzhanovsky (1976) がパンボルス族 (Pamborini) を設けているが,Moore (1966) は幼虫の形態からセダカオサムシに近いと主張している。我々の核 28S rRNA 遺伝子による解析では, オーストラリアオサムシ, チリオサムシ, セダカオサムシ族およびオサムシ族はそれぞれ独立の系統となる。分岐順はセダカオサムシ, オーストラリアオサムシ, チリオサムシ, Carabini オサムシ族 (Calosomina + Carabina) となったが, ブートストラップ 値が低く, ほぼ同時に分岐したと考えられる。従って, オーストラリアオサムシとチリオサムシは分類学的にはそれぞれ別族 (Tribe) を形成していると結論される。また, mtND5 遺伝子による解析においても, ほぼ同様な結果が得られているが, オーストラリアオサムシとチリオサムシは系統樹の根元で弱く組み, クラスターを形成していることは興味深い。この結果はオーストラリアオサムシとチリオサムシが祖先を共有していることを示唆している。mtND5 遺伝子による年代推定では, オーストラリアオサムシとチリオサムシの分岐はおよそ 6000 万年前であり, この年代は南米大陸, 南極, オーストラリア大陸が分かれた年代 (1 億 2000 万年前~ 6500 万年前) よりやや新しいが, mtND5 遺伝子の塩基置換は 15% 程度になると塩基置換が飽和状態に近づくので, 実際の分岐年代はこの計算値より古い可能性があると思われる。またオーストラリアオサムシとチリオサムシはそれぞれオーストラリア大陸と南米のチリにしか分布しておらず, 他のオサムシと全く姉妹関係を示さないため, ユーラシア大陸や北米から進入してきたとは考えにくい。したがって, オーストラリアオサムシとチリオサムシは南米大陸, 南極, オーストラリア大陸が陸続きの時代に共通祖先から大陸移動によって隔離されてそれぞれの分布域が成立したと考えられる。他のオサムシの起源と分布圏の成立に関しては, オサムシの上位グループ間の分岐年代を確実に測定した上で考えたい。いずれにせよ, 大陸移動の他に, 例えば島づたいなどによる進入経路も考えられる。
オサムシの一斉放散
オサムシ亜族 (Carabina) は形態的特徴 (♂交尾器を重視) から 2 群 (真正オサムシ群と多条オサムシ群) と 8 亜群に分類された。mt DNA の系統樹は, ほぼこの分類を支持したが, 各亜群はほぼ同時期に分岐, 放散しているので, 上位の群を廃し, 亜群を群に昇格した。現在, オサムシ亜族は9つの群に分けられており, 約 130 (亜) 属を含んでいる。その多くは中国大陸奥地の西南部に分布している。本報告で用いる種名, 亜種名は基本的には「世界のオサムシ大図鑑」に従った。また便宜上, 亜属名を属名として記述した。
オサムシ亜族に属する約 130 の (亜) 属の代表的な種類 (1,500 個体以上) の mtND5 遺伝子の塩基配列を比較して系統関係を調べたところ, これらの (亜) 属の多くは一斉に分岐したことが分かった (一斉放散) 。代表的な種類の mtND5 遺伝子の塩基配列を用いて作成した系統樹は図2である。この一斉放散は短期間の間に形態の異なる様々な種が爆発的に誕生したことを意味する。一斉放散の時期は約 5000-4000 万年前と推定され, インド大陸がユーラシア大陸に衝突し, ヒマラヤが隆起した時期とほぼ一致する。急激な地殻変動に伴う環境変化が, 短期間での形態多様化を誘発したとも考えられるが, オサムシ亜族の祖先が古インド亜大陸の北の一部にのってきてユーラシア大陸に到達し急速に分布を拡大する過程で放散したとも考えられる。オサムシ族の起源に関する更なる検討が必要である。いずれにせよ, 一斉放散後, それぞれの系統内でも, 中, 小規模の放散が起き, 現在のオサムシの種が出揃ったと思われる。
同じような一斉放散はカタビロオサムシ亜族 (Calosomina) やセダカオサムシ族 (Cychrini) でもみられる。中国,韓国,日本,アメリカ,ニカラグア,エクアドル,オーストラリアなどの国から採集した12種,17個体カタビロオサムシの mtND5 遺伝子の塩基配列を決定し,系統解析を行ったところ,これらのオサムシはほぼ同時に分かれ,オサムシ亜族 (Carabina) で見られたような一斉放散がカタビロオサムシ亜族でも起きたことが分かった。一斉放散の時期は Carabina 亜族と比べると, やや後の 約 3500-3000 万年前と推定された。
中, 小規模の放散はヨロイオサムシ群 (Procrustimorphi) や骨片オサムシ群(Digitulati) をはじめとするほとんどすべての群でみられ, 形態変化はヨロイオサムシ群でもっとも顕著, 骨片オサムシ群ではそれほどでない。中国系ヨロイオサムシは特に形態が多様化しているグループで, mtND5 遺伝子による系統解析の結果は図3に示されている。まず6系統:Shenocoptolabrus/Acoptolabrus/Coptolabrus/Cathaicus(1), Aristocarabus/Shunichiocarabus/Pagocarabus/Eccoptolabrus/Calocarabus/Neoplesius/Eocechenus(2),
Cephalornis(3), Damaster(4), Coptolabrodes(5), Acathaicus(6) がほぼ同時に分かれ, そのうちの1つ (2) は後ほどさらに4つの系統に一斉に分岐した。放散時期はそれぞれおよそ 3000 万年前と 2000 万年前であると推定された。
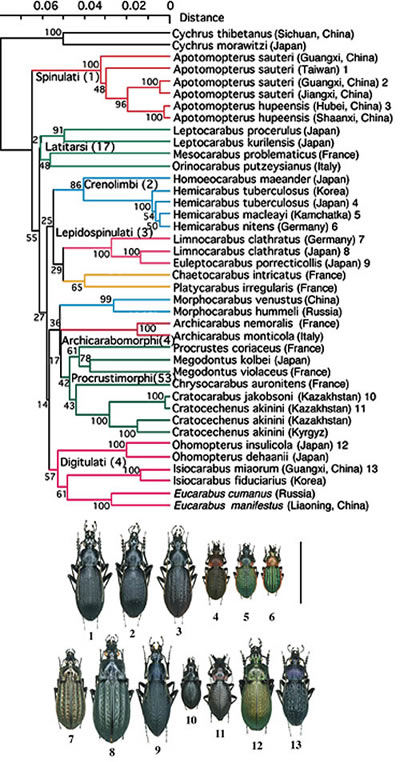 |
【図2:世界の代表的なオサムシ亜族(Carabina)の系統樹】
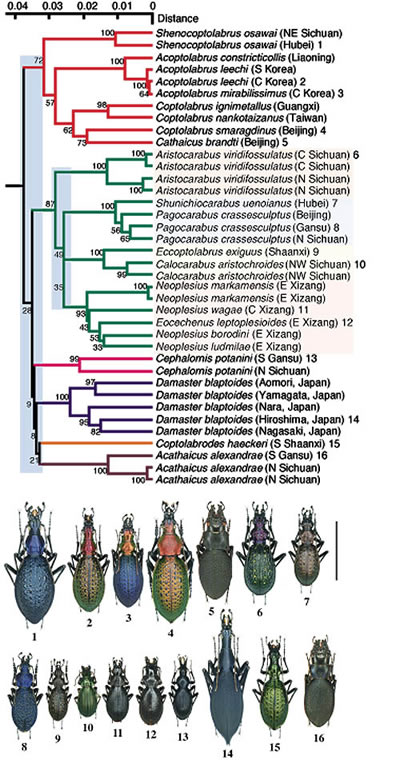 |
【図3:中国系ヨロイオサムシ(Procrustimorphi)の系統樹】
不連続形態進化
オサムシの進化過程において, ある形態種から, 別の形態種への転換もしばしば起きたと推定される。チベット高原から四川省にかけて分布するチベットオサムシ (Neoplesius spp.) (N)は単系統で, ~1,400万年前に一斉に放散しているが, 同所的に分布する頭胸部が著しく肥大したタカネオオズオサムシ (Eocechenus) (E)もこの中に入る。N→Eの形態変化が急速に起きたことを示唆している。このような頭胸部の肥大化は, サドマイマイカブリ (Damaster blaptoides capito) や, 韓国のオオズクビナガオサムシ (Acoptolabrus mirabilissimus), その他でもしばしばみられるが, 系統樹ではサドマイマイカブリは本土のコアオマイマイカブリ (D. b. babaianus) 中に埋没し, オオズクビナガは同所的に産するリーチホソクビナガ (A. leechi) とmt DNAでは全く区別できない。異様なまで巨大化した頭部を持つマンボオサムシ (Acathaicus) (中国四川省) に最も近縁とされる中国北部のコウガオサムシ(Cathaicus) とクギヌキオオズオサムシ (Eupachys) は, 全く形態の違うカブリモドキとクビナガオサムシとそれぞれ姉妹関係にある (図4) 。アキタクロナガオサムシ (Euleptocarabus porrecticollis) は, ~1,000万年前に本州 (多分中部) でマークオサムシ (Limnocarabus clathratus aquatilis) から急激な形態変化を伴って分化したと推定される。これらの例 (まだ外に多数ある) は, 非連続形態変化が, 時には大規模に, またある時には小規模に起きたことを示唆している。非連続な大変化 (explosive evolution) は, 動物の門の一斉出現 (カンブリアの大爆発) やアフリカのビクトリア湖およびその周辺の湖におけるカワスズメ (cichlids) などが有名で, オサムシの一斉放散に限った話ではない。
同じような形態変化は時に別系統に並行して起こることがあり, 平行進化 (parallel evolution) と呼ばれる。ここで日本特産のオオオサムシ亜属 (Ohomopterus) の系統関係を紹介するが, その前にこの亜属の起源について簡単に述べることにする。オオオサムシ亜属はアカガネオサムシ亜属 (Carabus), ツヤオサムシ亜属 (Eucarabus), タイリクオオオサムシ亜属 (Isiocarabus) の3亜属と共に, 骨片オサムシ群 (Digitulati) に属する。 骨片オサムシ群は雄交尾器の内袋腹面中央に指状片 digitulus (交尾片または骨片) を備えていることが最大の特徴で, ユーラシア大陸から北米大陸まで, 全北区にわたる広い分布域を持っている。世界各地から集められたこれらのオサムシのミトコンドリア ND5 遺伝子を解析し, 系統樹を作成した。この系統樹によると, 骨片オサムシ群は少なくとも6つの系統に分かれることが判った。(1) アカガネオサムシ (Carabus granulatus)/ホソアカガネオサムシ (C. vanvolxemi) /コブスジアカガネオサムシ (C. arvensis) ;(2) ツヤオサムシ亜属;(3) 北米のヒメホクベイアカガネオサムシ (C. limbatus) ;(4) granulatus 以外の中国産アカガネオサムシ各種;(5) タイリクオオオサムシ亜属;(6) 日本特産のオオオサムシ亜属。ツヤオサムシ亜属, タイリクオオオサムシ亜属とオオオサムシ亜属はそれぞれ単系統としてグループを形成したが, これまでアカガネオサムシ亜属として纏められたオサムシは少なくとも3つ (上記 1, 3, 4)の独立した系統に分かれていたことが示唆された。タイリクオオオサムシは形態的にオオオサムシに近縁で, オオオサムシの直接祖先として考えられていたが, 分析した中国南部と韓国済州島のタイリクオオオサムシは全く別系統であり, 日本のオオオサムシ亜属の起源とは思われない。
日本のオオオサムシ亜属は, 主として♂交尾器の骨片の形態からヒメオサムシ japonicus (J), オオオサムシ dehaanii (D), ヤコンオサムシ yaconinus (Y), アオオサムシ insulicola (I)の4タイプ (種群) に分けられ, それぞれの中には多数の種や亜種が設定されている。mtDNAの系統樹では, 形態の区分けのようにはならず, 地域的な8系統が認められ, 一つの系統に複数のタイプが混在する。我々はこの結果を, Ohomopterus の種分化の過程で, 各地域で同一タイプが独立に平行的に生じたと推定し, これを平行放散進化, その変化の様式をタイプスイッチングと名付けた。
タイプスイッチング説に関して, 色々な議論がある。私達もOhomopterus のミトコンドリア ND5 の系統樹がコンピューターの画面に現れたとき, これまでの形態によるものと余りにも違うことに驚いた。しかし, この系統樹を解釈しなければならない。タイプスイッチング説以外に, ミトコンドリア DNA の祖先的多型や交雑によるミトコンドリア DNA の水平移動などの可能性が考えられる。しかし, 地域的系統が綺麗に形成されていることからミトコンドリア DNA の祖先的多型はあり得ないと判断した。交雑によるミトコンドリア DNA の水平移動については部分的に多少あるかもしれないが, これだけではやはり得られたミトコンドリアの系統樹関係を説明することができない。例えば西日本のオオオサムシとヒメオサムシはミトコンドリア ND5 遺伝子の系統樹では四国, 九州・山陰, 山陽と3系統にほぼ同時に分かれた。交雑によるものと仮定すると, 多分それぞれの地域でミトコンドリアが全部どちかの方に置き換わっていると考えざるを得ない。たとえそうだとしても, 一つはオオオサムシのミトコンドリア, もう一つはヒメオサムシのミトコンドリアとした場合, 三つ目の系統のミトコンドリアの起源は不明になる。また対馬のヒメオサムシ, 隠岐の島のオキオサムシ, それから東日本のアオオサムシはすべてクロオサムシからミトコンドリアをもらってきたと考えなければならないが, 対馬と隠岐島にはクロオサムシが分布していない。更にこれらのことを考える前提条件として, ヒメオサムシとオオオサムシ, クロオサムシとアオオサムシ, それぞれ交雑可能でなければならない。このような交雑実験が必要であろう。
核 DNA の系統樹と, ミトコンドリアの系統樹との整合性を調べることは, 1つ有効な手段として考えられる。しかし核 DNA は一般的に進化速度が遅いのでこのオサムシグループの系統解析は困難と思われる。私達は核 DNA の中で最も進化速度が速いと思われる ITS 領域で系統解析をしてみた。Ohomopterus グループ全体の系統関係を語る信頼性の高い系統樹は得られなかったが, 部分的には Type switching 仮説へのサポーティングデータが得られている。核DNA ITS を約1kb の配列を比べたところ, 東日本のアオオサムシグループとミカワオサムシ (O. arrowianus) などを含む中部アオオサムシグループははっきり異なる。境界は糸魚川-静岡構造線の辺りで, ほぼミトコンドリア DNA のそれと一致する。またイワワキオサムシ (O. iwawakianus) とマヤサンオサムシ (O. maiyasanus) についても解析が進んでいる。イワワキオサムシは5角形の交尾器を持つヤコンオサムシグループで (Y-type) , マヤサンオサムシはアオオサムシグループである (I-type)。紀伊半島にマヤサンオサムシの亜種とされる, タキハラオサムシ (I) とシラガキオサムシ (I) がそれぞれイワワキオサムシ或いはキイオサムシ (イワワキオサムシの亜種) 分布域の中に点在分布している。両亜種 (タキハラ, シラガキ) の核 ITS 配列はイワワキオサムシ (Y) と殆ど同じで, マヤサンオサムシ (I) とは全く違う。もしこの個体は雑種であると仮定すると, マヤサンオサムシとイワワキオサムシ両タイプのITS が同じ個体に存在するはずだが, 検出されていない。イワワキオサムシ (Y) からタキハラオサムシ, シガラキオサムシ (I) への type switching の可能性が大きいと考えられる。
タイプスイッチングによる平行進化は, Ohomopterus に限った話ではなく, 他の分類群, 或いは上位の分類群でも見られる。その例としてクロナガオサムシ属 (Leptocarabus) をあげる。Leptocarabus は日本以外では主としてユーラシア大陸北部に広く分布するが, その中で朝鮮半島のセイシンコクロナガオサムシ (L. seishinensis) とチョウセンコクロナガオサムシ (L. semiopacus) は, 日本のコクロナガオサムシ (L. arboreus ) と形態的に極めて近縁とされ (Sタイプ) , 中国中南部に産するクロナガオサムシ (L. yokoae/L. marcilhaci) (図5: 2, 3)は日本のクロナガオサムシ, 特にキュウシュウクロナガオサムシ (L. kyushuensis) (図5: 1)にもっとも近縁とされている (Pタイプ) 。しかし, mt DNAと核28S rDNAの系統解析結果, Leptocarabus は3つの独立した系統に分かれることが分かった:1)中国中南部に産するクロナガオサムシ;2)1以外の大陸産クロナガオサムシ;3)日本産クロナガオサムシである。この結果は, Sタイプが別のタイプから大陸と日本で多分タイプスイッチングによって独立に, 平行して生じたことを示唆している。さらに驚いたことには, クロナガオサムシを全体のオサムシ亜族の種とともに系統解析を行ったところ, 中国中南部産のクロナガオサムシは全く形態的に遠縁のドウガネオサムシ属 (Rhigocarabus) に近いことが示唆された (図5) 。核 28S rDNA の解析でも全く同様な結果が得られた。中国中南部産のクロナガオサムシは, 他のクロナガオサムシと起源が異なり, ドウガネオサムシと祖先を共有した, 全くの別系統であることが判明された。このことは, オサムシの上位分類群でもタイプスイッチング機構が存在することを示唆している。
また本文の前にも述べたように, 異様なまで巨頭化した2種類のオサムシ, コウガオサムシ (Cathaicus brandti) とクギヌキオオズオサムシ (Eupachys glyptopterus) は形態的にもっとも近縁と思われていたが, mtND5 遺伝子の系統樹では, それぞれカブリモドキと大陸のクビナガオサムシとクラスターを形成している (図4) 。このような巨頭化した, 真っ黒のオサムシが美麗なオサムシの中でも群を抜くカブリモドキとクビナガオサムシと, それぞれ組むことには驚きを禁じえない。これも1種のタイプスイッチングとみてもいいと思われる。
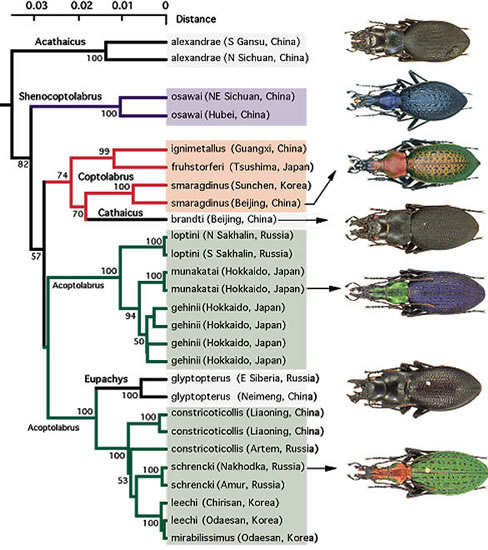 |
【図4:コウガオサムシ(Cathaicus)の系統関係】
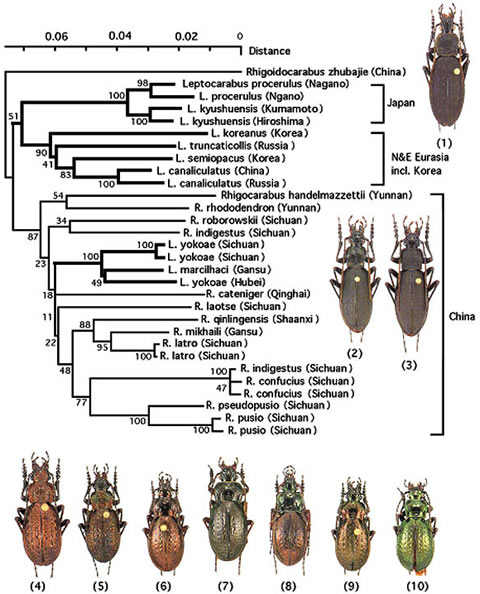 |
【図5:中国産クロナガオサムシ(Tomocarabus opaculus)の系統関係と産地図】
オサムシの進化における「動」と「静」
形態多様化が非連続的に起こるということは, 形態変化がほとんど起きない時期があることを意味している。ザウタートゲオサムシ (Apotomopterus sauteri) は, 中国と台湾に分布し, ごく軽微な形態差でいくつかの亜種に分けられているが, その分岐は古く, ~2,000万年前と計算される。マイマイカブリの近畿以西の3系統は形態的に区別できないがその分岐は~1,300万年前である。ヨーロッパに広く分布するクロツヤオサムシ (Phricocarabus glabratus) は形態的にほとんど区別できないが, mt DNAでみるとアルプスを境に2系統に分れ, その分岐はアルプス形成の~2,000万年前と計算される。同じことがダルマオサムシ (Tomocarabus convexus) , チョウセンヒメダルマオサムシ (T. fraterculus), ヒメクロオサムシ (T. opaculus) などでもみられる。ダルマオサムシ属 (Tomocarabus) の放散は前述した中国系ヨロイオサムシ (図3) とほぼ同時期 (3000万年前) であるが, 形態はほとんど変化していない。特に放散後の各系統内では, 形態の多様化は全く起きなかったと言っても過言ではない (図6)。非連続な形態変化を「動」とすれば, 上にあげたものを「静」と表現することができよう。例示した「静」の時期は, 実にオサムシ亜族の歴史の1/2~1/3の長期にわたっている。一般には, 地理的隔離が種形成の主因と云われるが, 果たしてそうであろうか。上に述べたことは, 隔離自身は顕著な形態変化の原因とならないことを示唆している。「生きた化石」としてあげられる他の生物の例は「静」の時期の長い例にすぎず, 例外的なものではない。進化は静と動の組合せによると考えられる。
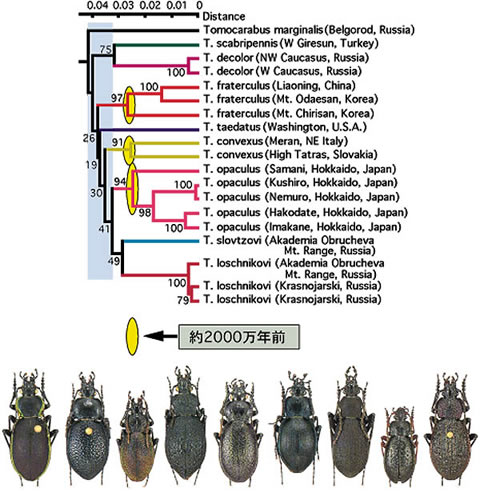 |
【図6:ダルマオサムシ(Tomocarabus)の系統関係】
日本のオサムシの起源
日本のオサムシは, すべて氷河期(約200万年前以後)に, 陸橋を通して日本列島に進入し, 列島内で分岐したと考えられている。しかし, mtND5遺伝子による系統解析の結果によれば, 列島への進入経路は一様ではなく, 大きく2つのグループに分かれることを示唆している。第1グループは, 約1,500万年前, 日本列島が大陸から分離した際, 祖先型が古日本列島に乗って入り, 列島内で分化したものであり, それぞれの国内での分岐は古い。第2グループはユーラシア大陸で分化し, 氷河期に陸橋によりサハリンまたは千島経由で北海道に入ったものと, 朝鮮半島から対馬に入ったものであり, 列島内での分岐はごく最近で, 大陸のものと区別がつかないほど近縁である。以上の推定は, 日本各地のサンプルと大陸沿海州, サハリン, カムチャツカ半島, 朝鮮半島のそれぞれの同種または近似種との進化距離を計算することにより導かれたものである (図 7, 図8) 。
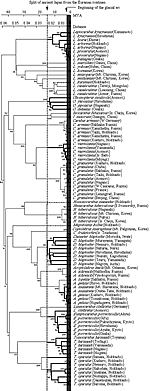 |
【図7:日本産オサムシ、および関連した大陸のオサムシの系統樹】
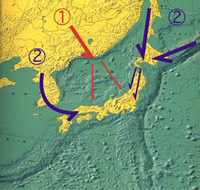 |
【図8:日本産オサムシ進入経路】
1:古日本列島にのって進入 2:氷河期の陸橋にのって進入
オオオサムシ亜属 (Ohomopterus) も日本特産のオサムシで, 15種に分類されている。この亜属内の分岐はマイマイカブリより少し新しく, おそらく日本列島が大陸から分離したやや後に分岐を開始したと考えられる。この亜属の祖先型は最初は古日本全域には分布しておらず, 西日本弧に乗って大陸からきた可能性が高い。この亜属は前述のように中国大陸, 朝鮮半島の北部, 済州島に分布するタイリクオオオサムシ亜属 (Isiocarabus) と形態が似ており, それから由来したと考えられているが, mtND5系統樹では, 両者は全く類縁関係を示さず, 直接共通祖先から分かれたもの同志ではない。
アキタクロナガオサムシ (Euleptocarabus porrecticollis) も日本特産亜属で, 本州にのみ分布している。mtND5系統樹では, アキタクロナガオサムシは日本のマークオサムシ (Limnocarabus clathratus aquatilis) と最も近縁で, ドイツのマークオサムシ (Limnocarabus clathratus clathratus) は外群になる。アキタクロナガオサムシの中では, 中部・近畿のものは比較的分岐が古く, 関東以北のものはごく最近分布を拡大した集団である。これらの系統関係と進化距離から, アキタクロナガオサムシは約1100万年前, 日本列島内 (多分中部地方) でマークオサムシから分化したと推測される。
日本のクロナガオサムシ類 (Leptocarabus) はクロナガオサムシグループと, コクロナガオサムシグループに分けられている。朝鮮半島にもクロナガオサムシグループとコクロナガオサムシグループが分布しており, クロナガオサムシ同志, コクロナガオサムシ同志がそれぞれ近縁とされている。しかし, mtND5と核 28S rRNA 遺伝子の系統解析では, 日本のクロナガオサムシと日本のコクロナガオサムシ, 韓国のクロナガオサムシと韓国のコクロナガオサムシが, それぞれクラスターを形成し近縁である。チシマオサムシを除く日本のすべての Leptocarabus (s.lat.) は単系統で, 日本列島内で種分化を起こしたと考えられ, その直接姉妹関係にある種は朝鮮半島を含む大陸からは発見されていない。したがって, 日本のクロナガオサムシ類の起源を朝鮮半島に求めるのは, 誤である。日本のクロナガオサムシの祖先型は大陸における古日本域に分布していたものであり, 約1,200万年前, キュウシュウクロナガオサムシと他のクロナガオサムシ/コクロナガオサムシが分岐, 600万年前以降, オオオサムシ亜属に似た経路で日本列島内で分布を拡げ, 多様化したと考えられる。
ホソアカガネオサムシ (Carabus vanvolxemi) , ヒメクロオサムシ (Tomocarabus opaculus) , ホソヒメクロオサムシ (T. harmandi) 3種についてはユーラシア大陸で同種, または近似種を見出すことができない。例えば, ホソアカガネオサムシは, 日本国内の中部から東北にかけて分布する固有種だが, これと近似かもしれないと考えていた中国のアカガネオサムシ類はすべて ホソアカガネオサムシとは全く別クラスターに入るし, 朝鮮半島にもホソアカガネオサムシの系統に入るものはいない。一つの考え方としては, Carabus (s. str.)放散の過程で, 大陸で約2,000万年前, アカガネオサムシと分れたホソアカガネオサムシの祖先型が大陸の東北周辺部に局所的に隔離され, 列島形成時に東日本弧に乗ってきた。多島化の時期に残された島に残存していた祖先型が, 続いて起きた陸化により, 近年その分布を拡大した, というものである。ヒメクロオサムシとホソヒメクロオサムシについても, 基本的にはホソアカガネオサムシと同じように考えられる。ヒメクロオサムシの分岐開始は, マイマイカブリよりも古く, 約2,000万年前, 北海道での分岐開始が約1,300万年前となる。おそらく大陸の古北海道域で既に2系統(A, B)に分かれていたのであろう。本州のヒメクロオサムシ (チョウカイヒメクロオサムシ) は上の2系統の一つ, Bに属し, 北海道のヒメクロでは様似町のものだけがこれに含まれるが, B系統内での分岐開始はごく最近である。北海道のB系統のヒメクロオサムシが本州に入り, 東北地方に分布を急速に拡げたか, 逆に本州のどこかに局在していた B 系統のヒメクロオサムシが最近急速に分布域を広げ, 下北半島で北海道にと推定される (図 9) 。
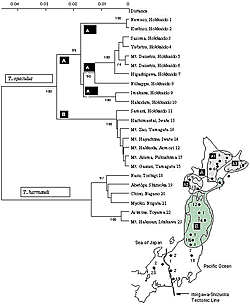 |
【図9:ヒメクロオサムシ(Tomocarabus opaculus)の系統関係と産地図】
第2グループは氷期に陸橋により日本列島に進入したもので, アカガネオサムシ (Carabus granulatus) , コブスジアカガネオサムシ (C. arvensis) , チシマオサムシ (Aulonocarabus kurilensis) , セアカオサムシ (Hemicarabus tuberculosus) , ツシマカブリモドキ (Coptolabrus fruhstorferi) などが含まれる。これらのオサムシの特徴としては, 日本列島内の各地のサンプル間のmtDNAの差が僅小で, 互いに近く, さらに, サハリン (または朝鮮半島;ツシマカブリモドキの場合) の同種または近似種とも mtDNA の差がほとんどないことがあげられる。ツシマカブリモドキは韓国南部のアオカブリモドキと極めて近いので, 朝鮮半島経由で対馬に入ったとみてよい。セアカオサムシは韓国, 沿海州, カムチャツカ半島のもののどれとも近いので, 北から進入したのか, 南から入ったのか, またはその両方から入ったと考えられる。他のものはサハリンのものと近く, おそらくサハリン経由で北海道に入ったと思われる。
「オオルリオサムシ (Acoptolabrus gehinii) は, 北海道にしか生息しておらず, 最終氷期にサハリン経由で進入したのであろう」とか, 「オオルリオサムシ(オシマルリオサムシを含む)はその形態の類似から大陸のクビナガオサムシに近いのではないか」との説があるが, 客観的な証拠はない。ND5 遺伝子から推定されたクビナガオサムシの系統関係によれば, クビナガオサムシ亜属 (Acoptolabrus) は, 大陸に分布するクビナガオサムシグループ(シュレンククビナガオサムシ, ホソクビナガオサムシ, リーチホソクビナガオサムシ, オオズクビナガオサムシ)と, カラフトクビナガオサムシ (A. lopatini) , オオルリオサムシ (A. gehinii) からなるグループの2つにはっきりと分けられる。クビナガオサムシが2グループに分かれたのは2,000万年前である。カラフトクビナガオサムシ, オオルリオサムシのグループの祖先種は日本列島形成に関わる地殻変動のかなり早い時期, おそらくサハリンと北海道の起源となる島が形成されたときに大陸のクビナガオサムシと隔離されたのであろう。一方これらの両グループ内での現存種への放散は, 両グループへの分化のずっと後になってから起こっており, およそ400万年前以降と考えられる。北海道のオオルリオサムシともっとも近縁な種は, カラフトクビナガオサムシであり, 大陸の種はいずれも遠縁である。ただし, その分化の時期は, 最終氷期等のごく最近のことではなく, 第三期にまでさかのぼるようである。以上の事実からオオルリオサムシは, 第1, 第2ブループとは多少異なるカテゴリーに属するようであるが, 北方地域の地史には不明な点が多いので今後の課題としたい。
セスジアカガネオサムシ (Homoeocarabus maeander) とアイヌキンオサムシ (Megodontus kolbei) については, 大陸やサハリンの資料がないか, または不足で, 第1グループに属するのか, 第2グループに入るのか不明である。
中国のオサムシ調査
1999年 5月 27日より 16日間, BRH と中国科学院成都分院が共同で, DNA 分析用サンプルの採集を目的とした第3次中国調査が行われた。今回調査したのは四川省北東端から陜西省南西部にかけての地域で, 米倉山と呼ばれる山塊ならびに漢中北部にある紫柏山一帯である。この調査では今世紀最大の発見とも言われる Shenocoptolabrus osawai が多数得られ, カタビロオサムシとセダカオサムシを含むやく 14種, 250頭以上採集することができた。これらのオサムシの解析結果も本報告中に含められている。
おわりに
オサムシ研究プロジェクトは1994 年に開始して以来,6年が経過した。この間, オサムシの一斉放散, 平行放散進化, 地史との関連, 不連続的形態変化, 及び進化における「動」と「静」など数々のことが解明され, 生物の進化, 或いは多様化の原理を探る上でも極めて重要な知見が得られたと考える。オサムシ研究プロジェクトは今年度で終了することになったが, この研究から発した昆虫系統解析についての研究は, 当プロジェクトの成功に触発されて最近日本中に広がり盛んに行われるようになった。このことからも本研究の成果の日本昆虫系統・分類学の発展に対する高い貢献度をうかがわせる。
学会発表など
総説・紹介
大澤省三 (1998)
小さな虫から日本列島の誕生が分かる. 三井グラフ, 112: 28-29.
大澤省三 「昆虫と自然」1999年2月増大号
特集:DNA で探る昆虫の系統進化
「はじめに-分子系統とは何か、何が分かるか」2-3.
大澤省三 「昆虫と自然」1999年2月増大号
特集:DNA で探る昆虫の系統進化
「オサムシにおける形態多様化の道のり」15-19.
大澤省三、池田清彦、柏原精一
「現代進化論を虫目敢する。オサムシから見えてきた生き物の「奥」の深さ。」(対談)
朝日新聞社 SCIaS , (98/1/02): 58-61.
小関治男、大澤省三
「分子生物学の黎明期」(対談)
朝日新聞社「DNA の謎に挑む」渡辺政隆著、 (1998): 303-317.
蘇 智慧、金 衝坤 「昆虫と自然」1999年2月増大号
特集:DNA で探る昆虫の系統進化
「オサムシの来た道を DNA で辿る」4-10.
蘇 智慧、金 衝坤(1999)オサムシの来た道を DNA で辿る
特集:DNA で探る昆虫の系統進化
「昆虫と自然 (1999年2月増大号)」 34: 4-10.
大澤省三 (1999)はじめに 分子系統とは何か、何が分かるか
特集:DNA で探る昆虫の系統進化
「昆虫と自然 (1999年2月増大号)」 34: 2-3.
蘇 智慧(2000)
ミトコンドリア DNA から見たオサムシの多様化 SHINKA(印刷中)
蘇 智慧(2000)
DNA でたどるオサムシの多様化 月刊海洋(印刷中)
大澤省三(2000)
完成ま近 DNA による世界のオサムシ系統樹 サイアス 2月号 6-11.
学会・シンポジウムなどの口頭発表
大澤省三・蘇 智慧ほか
大澤省三
オサムシの来た道行く道
広島虫の会 第36回総会 1998.3.8. (広島市舟入高校,広島)
大澤省三
DNAで辿るオサムシの進化
第13回「大学と科学」公開シンポジウム「遺伝子で生物の進化を考える」
1998.10.13-14(東京)
大澤省三
オサムシの分子系統から見た進化の様式
「生物多様性」懇談会 1998.12.22.(京都)
大澤省三
DNA による系統解析とは何か、何がわかるか
日本鞘翅学会第 11 回「甲虫の DNA による系統解析分科会」1998.11.14-15(松山)
蘇 智慧、井村有希、大澤省三
ミヤマオサムシ (Oreocarabus) 亜属の分子系統関係
第42回日本応用動物昆虫学会大会 1998.3.31-4.2(名古屋)
Su, Z.-H.
Parallel evolution and rapid morphological differentiation in the Carabinae ground beetles (Coleoptera, Carabidae) based on mitochondrial DNA sequences.
CAST 3rd Academic Conference of Young Scientists 1998.8.20-22 (Beijing)
蘇 智慧、井村有希、大澤省三
ミトコンドリア ND5 遺伝子塩基配列比較による骨片オサムシ亜群 (Digitulati) の系統関係
日本昆虫学会第58回大会 1998.10.2-4(彦根)
蘇 智慧、井村有希、大澤省三
中国四川省産オサムシの生息環境と系統関係について
日本鞘翅学会第 11 回 1998.11.14-15(松山)
蘇 智慧
オサムシの DNA 解析の現状
日本鞘翅学会第 11 回「甲虫の DNA による系統解析分科会」1998.11.14-15(松山)
金 衝坤、蘇 智慧、井村有希、大澤省三
トゲオサムシ亜群 Spinulati の分子系統
第42回日本応用動物昆虫学会大会 1998.3.31-4.2(名古屋)
金 衝坤、蘇 智慧、岡本宗裕、井村有希、大澤省三
ヨロイオサムシ亜群 Procrustimorphi の分子系統
日本昆虫学会第58回大会 1998.10.2-4(彦根)
Kim, C.-G., Su, Z.-H. and Osawa, S.
Phylogeny and evolution of the Apotomopterus ground beetles (Coleoptera, Carabidae) based on morphology and mitochondrial DNA sequences.
韓国遺伝学会 1998.10.30-31(高麗大学,ソウル)
金 衝坤
分子系統学の最前線
韓国土壌動物学会特別講演 1998.11.7(全北大学,韓国)
金 衝坤、蘇 智慧、岡本宗裕、白 種哲、井村有希、大澤省三
韓国産オサムシのミトコンドリア DNA による系統解析
日本鞘翅学会第 11 回 1998.11.14-15(松山)
金 衝坤
DNA と形態
日本昆虫学会第58回大会 1998.10.2-4(彦根)
斉藤秀生、金 衝坤、蘇 智慧、大澤省三
ミトコンドリア ND5 遺伝子から見たキタカブリとエゾマイマイカブリの分布境界について
日本鞘翅学会第 11 回 1998.11.14-15(松山)
蘇 智慧
オサムシの分子系統と地史との関連
東京大学海洋研究所共同利用シンポジウム「分子海洋学――分子生態学と海洋学の接点」1999.3.2-3(東京)
蘇 智慧、金 衝坤、井村有希、大澤省三
オサムシ亜科 (Carabinae) の高次分類群の分子系統関係
日本応用動物昆虫学会第 43 回大会 1999.4.2-4(那覇)
金 衝坤、蘇 智慧、冨永 修、大澤省三
分子系統樹からみたクロナガオサムシ亜属 (Leptocarabus) の起源と種分化
日本応用動物昆虫学会第 43 回大会 1999.4.2-4(那覇)
大澤省三
DNAで辿るオサムシの進化
国立遺伝学研究所開所 50 周年記念講演 1999.5.1(三島・国立遺伝学研究所)
蘇 智慧
ミトコンドリア DNA から見たオサムシの多様化
第24回進化学研究会例会「昆虫の分子系統学の最前線:DNAからなにがわかるか」
1999.9.4(京都)
蘇 智慧、金 衝坤、井村有希、冨永修、岡本宗裕、大澤省三
ミトコンドリア DNA から見たオサムシの進化様式
日本遺伝学会第 71 回大会 1999.9.24-26 (広島)
金 衝坤、蘇 智慧、冨永修、井村有希、大澤省三
オサムシの平行進化 (Parallel evolution)
日本遺伝学会第 71 回大会 1999.9.24-26 (広島)
蘇 智慧、金 衝坤、井村有希、冨永 修、岡本宗裕、大澤省三
オサムシの非連続的進化
日本鞘翅学会第 12 回 1999.11.13-14(豊橋)
冨永 修、蘇 智慧、金 衝坤、大澤省三、岡本宗裕、井村有希
日本列島におけるオサムシ相形成過程の推定
日本鞘翅学会第 12 回 1999.11.13-14(豊橋)
井村有希、金 衝坤、蘇 智慧、大澤省三
分類学的位置の決定が困難であったいくつかのオサムシの分子系統
(おもに西ヨーロッパ、北アフリカ産の種を中心として)
日本鞘翅学会第 12 回 1999.11.13-14(豊橋)
斉藤秀生、斉藤明子、金 衝坤、蘇 智慧
チャイロヒメハナカミキリ種群数種の類縁関係について
日本鞘翅学会第 12 回 1999.11.13-14(豊橋)
蘇 智慧、冨永 修、金 衝坤、大澤省三
ヒメクロオサムシの分子系統
日本鞘翅学会第 12 回「 DNA 分科会」1999.11.13-14(豊橋)
金 衝坤、蘇 智慧、岡本宗裕、井村有希、大澤省三
ヨロイオサムシ群の分子系統
日本鞘翅学会第 12 回「 DNA 分科会」1999.11.13-14(豊橋)
大澤省三
分子系統と形態分類の融合で進化を探る
日本動物分類学関連学会連合設立記念シンポジウム 2000.1.8(東京、国立科学博物館分館)
講演・会議参加
大澤省三
広島大学理学部生物科学セミナー「オサムシの来た道」(1998.3.9 広島大学、広島)
大澤省三
分子遺伝学・分子進化学(講義)(1998.6.9-11 信州大学理学部、松本)
大澤省三
平成10年高等学校「理科」教養研修 B (生物)「オサムシの進化(講義)」
(1998.10.8 大阪府教育センター、大阪)
大澤省三
平成10年高等学校「理科」教養研修 B (生物)「遺伝暗号の進化とオサムシの進化(講義)」
(1998.12.9 大阪府教育センター、大阪)
金 衝坤
韓国農業科学技術院(韓国水原)セミナー「全世界のオサムシの分子系統解析」
(1998.11.2 韓国)
金 衝坤
慶北大学セミナー「ミトコンドリア DNA から見た韓国産オサムシの系統的位置」
(1998.11.4 韓国大邱)
論文リスト
1) Su, Z.-H., Ohama, T., Okada, T.S., Nakamura, K., Ishikawa, R. and Osawa, S. (1996)
Phylogenetic relationships and evolution of the Japanese Carabinaeground beetles based on mitochondrial ND5 gene sequences. J. Mol. Evol. 42, 124-129.
2) Su, Z.-H., Ohama, T., Okada, T.S., Nakamura, K., Ishikawa, R. and Osawa, S. (1996)
Geography-linked phylogeny of the Damaster ground beetles inferred from mitochondrial ND5 gene sequences. J. Mol. Evol. 42, 130-134.
3) u, Z.-H., Okada, T.S., Osawa, S., David, B., Dommergues, J.-L. and Magniez, F. (1996)
Radiation of several Carabina groups (Coleoptera,Carabidae) inferred from the mitochondrial ND5 gene sequences. Elytra,Tokyo 24, 175-179.
4) Su, Z.-H., Tominaga, O., Ohama, T., Kajiwara, E., Ishikawa, R. Okada, T.S., Nakamura, K. and Osawa, S. (1996)
Parallel evolution in radiation ofOhomopterus ground beetles inferred from mitochondrial ND5 gene sequences. J. Mol. Evol. 43, 662-671.
5) Imura, Y., Su, Z.-H. and Osawa, S. (1997)
Morphology and Molecular Phylogeny of Some Tibetan Ground Beetles Belonging to the Subgenera Neoplesius and Eocechenus (Coleoptera, Carabidae). Elytra, Tokyo, 25 : 231-245.
6) Osawa, S., Su, Z.-H., Kim, C.-G., Okamoto, M., Tominaga, O. and Imura, Y. (1999)
Evolution of the Carabid ground beetles. Adv. Biophys., 36: 65-106.
7) Su, Z.- H., Tominaga, O., Okamoto, M. and Osawa, S. (1998)
Origin and diversification of hindwingless Damaster ground beetles within the Japanese islands as deduced from mitochondrial ND5 gene sequences (Coleoptera, Carabidae). Mol. Biol. Evol., 15: 1026-1039
8) Kim, C.-G., Tominaga, O., Su, Z.-H. and Osawa, S. (1999)
Origin and diversification of Euleptocarabus porrecticollis (Coleoptera, Carabidae) in the Japanese islands inferred from mitochondrial ND5 gene sequences. Mol. Phylogenet. Evol., (in press).
9) Imura, Y., Zhou, H.-Z., Okamoto, M., Su, Z.-H. and Osawa, S. (1998)
Phylogenetic relationships of some Chinese ground beetles belonging to the subgenera Neoplesius, Pagocarabus and Aristocarabus (Coleoptera, Carabidae) based on mitochondrial ND5 gene sequences. Elytra, 26: 1-7.
10) Imura, Y., Su, Z.-H. and Osawa, S. (1998)
Some cychrini species (Coleoptera, Carabidae) from central Sichuan, China: descriptions of two new species and evolutionary considerations. Elytra, 26: 9-16
11) Imura, Y., Kim, C.-G., Su, Z.-H. and Osawa, S. (1998)
An attempt at the higher classification of the Carabina (Coleoptera, Carabidae) based on morphology and molecular phylogeny, with special reference to Apotomopterus, Limnocarabus and Euleptocarabus. Elytra, Tokyo, 26: 17-35.
12) Imura, Y., Su, Z.-H., Kim, C.-G. and Osawa, S. (1998)
Reorganization of the Oreocarabus complex (Coleoptera, Carabidae) based on endophallic morphology and molecular phylogeny. Elytra, Tokyo, 26: 223-248.
13) Imura, Y. and Su, Z.-H. (1998)
Two new subspecies of the genus Carabus (s. lat.) (Coleoptera, Carabidae) from northern Sichuan, China. Elytra, Tokyo, 26: 249-255.
14) 澤野十蔵,伊谷 結,蘇 智慧,大澤省三 (1998)
ミヤジマトンボとその原亜種のミトコンドリアDNA の比較. Tombo, (in press).
15) Osawa, S., Su, Z.-H., Kim, C.-G., Okamoto, M., Tominaga, O. and Imura, Y. (1999)
Evolution of the carabid ground beetles.
Adv. Biophys. 36: 65-106.
16) Kim, C.-G., Tominaga, O., Su, Z.-H. and Osawa, S. (1999)
Origin and diversification of Euleptocarabus porrecticollis (Coleoptera, Carabidae) in the Japanese islands inferred from mitochondrial ND5 gene sequences.
Mol. Phylogenet. Evol. 13: 440-444.
17) 井村有希・周 紅章・蘇 智慧 (1999)
中国湖北省神農架から発見された顕著なヨロイオサムシの1新種。
月刊むし No. 341: 2-5.
18) Kim, C.-G., Saito, S., Su, Z.-H., Tominaga, O. and Osawa, S. (1999)
Distributional boundaries of the geographic races of Damaster blaptoides (Coleoptera, Carabidae) in northeastern Japan as deduced from mitochondrial ND5 gene sequences.
Elytra, Tokyo 27: 635-641.
19) Kim, C.-G., Su, Z.-H., Imura, Y,. and Osawa, S. (1999)
Origin and evolution of the Apotomopterus ground beetles (Coleoptera, Carabidae) as deduced from mitochondrial ND5 gene sequences.
Elytra, Tokyo 27: 643-649.
20) Imura, Y., Zhou, H.-Z., Su, Z.-H. (1999)
A new species of Apotomopterus and a new subspecies of Cychrus (Coleoptera, Carabidae) from China.
Elytra, Tokyo 27: 651-654.
21) Kim, C.-G., Zhou, H.-Z., Imura, Y., Tominaga, O., Su, Z.-H. and Osawa, S. (2000)
Pattern of morphological diversification in the Leptocarabus ground beetles (Coleoptera, Carabidae) as deduced from mitochondrial ND5 gene sequences.
Mol. Biol. Evol. 17: (in press).
22) Tominaga, O., Su, Z.-H., Kim, C.-G., Okamoto, M., Imura, Y. and Osawa, S. (2000)
Formation of the Japanese Carabina fauna inferred from a phylogenetic tree of mitochondrial ND5 gene sequences (Coleoptera, Carabidae). J. Mol. Evol. (in press).