



ラジオ少年
小さな頃は普通の子供でした。今は全くしないけど、当時は野球少年だったり短距離も速かったりとスポーツは得意でした。中学生の終わりに急に背が伸びて、バランスが悪くなったのかな、その後運動はあまりしていませんね。いつも後ろの方に座っている生徒で、授業は全然聞いていませんでした。とにかく数学が面白くて、ピタゴラスの定理には証明法が100あると聞いて、全部自分でできないだろうかと試しました。100まではいかなかったけど、あれこれ考えるのは楽しかったですね。
それにラジオ少年でした。今と違って真空管の時代で、セットになっているわけではなくて、部品を一つ一つ秋葉原まで買いに行っていました。中学2年生の時にスピーカーを改良しようとして母の腰巻きを使ってえらく怒られましたね。良い音を出すためにどうしても柔らかいフランネルの布が欲しかったのです(笑)。3年生になって、トランスを買いに直接工場を訪ねたら、中学生でそこまでやるのは偉いって、工場長さんが工場を案内してくれました。きっとその時とても面白かったのでしょうね、「ここに勤めたい」と言ったみたい。工場から、学校に連絡があり、親が呼び出され、高校に進学しなさいと叱られました。ですから、何かになりたいと思っていたわけでもなくて、わりあいのんびりしていましたね。
高校は都立両国高校です。進学校でね、成績は中ぐらいでしたかね。相変わらず数学に興味があって、大学の教科書を自分で勉強していました。その時に数学は面白いけれど専門の数学者になるのは無理かなと思ったことを覚えています。私は数学と物理はよくできたけど、あとはさっぱり。当時の両国高校は東大に合格させる人数を誇っているようなところがあって、進路指導で、お前のようなオールラウンドでないタイプの学生は高得点を必要とする東大の理数系には受からないよ、生物系なら受かるかも知れないと言われました。でも自分は生物には全く興味がなかったので、物理系を受験したのですが、本当に落ちちゃった(笑)。だから、早稲田大学の理工学部に行きました。
大学でも熱心に授業を受ける学生ではありませんでした。この先生は聞こうと決めた4人の先生の授業はきちんと聞いたけど後は図書館で理論物理の勉強をしていました。数学は高校の時に大学レベルの本を読んでどういう学問かを知っていたけれど、理論物理は大学に入って初めて出会った学問でしたから、新しいものに触れた喜びで、夢中で勉強しました。私の学生時代を思い返すと、今の学生は真面目すぎますね。つまり、授業によく出て、単位をとることに熱心すぎる。出欠をきちんと取ってくれって言われたりしますからね。そんなことより、自分が夢中になることを探すことが大事だと思います。

大学に入って初めて出会った学問「理論物理」。夢中で勉強した思い出の本。
研究に魅了される
4年生での卒論のための研究室は、素粒子物理。宇宙線を解析する仕事で、実測データをモデルに当てはめて理論を導くことを行ったのです。初めての体験にとても興奮し、引き込まれました。理論物理が好きだったので、それを勉強するのを面白がっていたのだけれど、ある視点からデータを分析するという面白さに魅了されてしまった。勉強ではなくて研究というものがあるのだということを実感したのです。私の卒業研究は論文にしなさいと教授が勧めてくれる程度にはまとまったので、ますます研究が面白くなりました。まだ英語は無理だからということで、日本語でしたが論文を書きました。
最初の研究がうまくいったこともあって、どうしても研究を続けたかったのですが、実は就職が決まっていた。仲間と2年生の春休みに西日本会社見学巡りをしていい会社だなと思っていた東洋レーヨンを受けたら、すんなり合格してしまったのです。でも、やはり研究を続けたいと思って東京教育大学の朝永振一郎先生の研究室を受験しました。同級生のすごく優秀なヤツも同じ研究室を受験していて、一緒に受かったんです。ただ、朝永先生は忙しくて直接指導してもらえないと指導教授から言われていたので少し悩みました。ちょうどその頃、湯川秀樹先生が素粒子の理論物理学者は日本に10人いれば十分で、これからの若い人たちは生物物理で活躍して欲しいということを『自然』という雑誌だったと思うんだけど、そこにお書きになっていたのです。友人には勝てないだろうという気持ちも少し手伝って、早稲田の新しくできた生物物理の講座に進学しました。この時、私には学問の将来が見えていたわけではないけれど、ここでの選択が私の今を決めていますね。
生物物理学の勃興
大学院に進学した頃、京都大学・名古屋大学・早稲田大学に新しく生物物理の講座が開設されたのです。優秀な素粒子物理の研究者が生物分野に転向して研究を始めていました。1960年代の生物物理学者のなかには、量子力学で生命現象を理解しようという量子生物学を目指していた人もいた。世界的にもそういう傾向がありました。1940年代に、シュレディンガーなどが、生命現象への関心を示したところからの動きで、一つは分子生物学へ、一方では量子生物学へと動いていたのです。フランスのプルマン博士、スウェーデンのレブディン博士、それから名大の右衛門佐(よもさ)重雄先生が中心でした。量子生物学は、例えば酵素の活性中心での反応を電子状態で把握しようというような研究でしたね。
その頃はコンピュータが研究に使えるようになり始めた時でしたので、私はコンピュータを使って量子力学的にタンパク質、DNAの電子状態を計算しようと試み始めました。当時はそんな計算ができるコンピュータはIBMにしかなくて、高い使用料を払って使わせてもらっていました。名古屋から電話で右衛門佐先生が、「こんな計算をしてみたらどうだろう」、などと言ってこられるのだけれど、こちらは学生で研究費など採れるわけがない。それでも計算をしてみたかったので、同じようにIBMにコンピュータを借りている建設関連の人と仲良くなって、アルバイトでダムの強度計算プログラムを書き、その代り少しコンピュータを使わせてもらうなど、学生なりの工夫をしたものです。そんな苦労はありましたが、新しい分野でしたから、かなり活発に研究ができました。
ところが、博士課程の途中から私は量子生物学に疑問を感じ始めたのです。そんな時、京都大学の基礎物理学研究所(基研)に松田博嗣先生が呼んで下さった。その頃の基研は、若い人の修行の場として半年ほど受け入れてくれる制度があったのです。銀閣寺の前の屋台で飲みながら、しょっちゅう皆で徹夜で議論しましたね。あの経験は素晴らしいものです。当時の基研の所長は湯川秀樹先生。若者が先生と議論できるサロンがありました。そこで、私は量子生物学に疑問を感じながらも、DNAに光を当てて、旋光分散を利用して、塩基配列の情報を得る方法を考えていたのです。旋光分散理論で塩基配列の情報が得られるのではないか、という話を湯川先生にしたら、先生が大いに気に入ってくれましてね。後でわかったのだけれど、湯川先生はそういう大きい話がすごく好きだったらしい。会うたびにあれはどうなったんだと聞かれるのは励みになりました。その後、アメリカの物性物理学の大御所モットー博士がDNAの塩基配列の研究に興味をもち、光ではなくて、熱で配列情報を得る方法を考えているらしいという話が聞こえてきました。相手は大御所ですから勝てないなぁとがっかりしていました。その時、湯川先生が、「研究ではそういうことはよくあるよ」と言葉をかけて下さったことをよく覚えています。
量子生物学から離れる
量子生物学には疑問を持ちながらも学位を取得して、名古屋大学の右衛門佐先生の研究室の助手になりました。先生方はすでに物理で専門基盤を持ち、生物が面白そうだからと研究している。その姿を見ると、自分は何を基盤とするのだと焦りや疑問を感じました。また、生命現象の現場を捉えようと思ったら、電子ではなく酵素やDNAといった分子が役者として演じている舞台ではないだろうかと思ったのです。これを確かめるには生物の中に一度どっぷり漬からなければいけないと思って、東大の野田春彦先生のところに1年間お世話になりました。でも生化学は自分には向いていないようでした。
そんな時、私を京大の基研に呼んで下さった松田博嗣先生が九州大学で教授になられて、「新しく数理生物学を始めるのだけれど、一緒にやらないか」と私を助教授で誘って下さった。数理生物学というのは、数学で生命現象を解明しようという研究分野です。具体的には松田先生は集団遺伝学を研究対象に考えておられたので、私は分子(DNA)から進化の研究に取り組もうと思い、73年に九州大学に移りました。
分子進化学へ
私が大学院生の時に塩基配列の情報を得る方法を考えていたことはお話しましたが、この頃にDNAを切断する制限酵素があることを知りました。これで、生化学的にDNAの塩基配列が決まる時代が来るに違いない。生物の進化の情報はDNAの変化から得られるはずだから、その情報をコンピュータを使って得ようと思いました。生化学には向いていないことがわかっていたので、自分で塩基配列を決めるのではなく、塩基配列を解析するコンピュータ・プログラムを作ろうと考えたのです。RNAファージなどですでに塩基配列が決まっていたものもあったので、それをあーでもない、こーでもないといじくり回していました。3年~5年試行錯誤が続きましたねえ。この時ありがたいことに、松田先生は私をほっておいて下さった。これは助かりました。うるさくいろいろ言われていたらどうなっていたかな。論文が全然でなかったのですから。タンパク質のアミノ酸配列は少しわかっていたのでそれを使って進化の研究をしましたが、コンピュータによる塩基配列の解析方法を探ることに主力を注いでいました。
1976年サンガーが、φX174ファージのDNA塩基配列を決めました。その中に、オーバーラップ遺伝子があることがわかったのです。オーバーラップ遺伝子では一つの塩基配列が2つの異なる読み枠で翻訳されて2つのタンパク質を作るために読まれるので、塩基が変化すると、通常の遺伝子よりも影響が大きいので、DNAが変化できないのではないかといわれていました。しかし実際に私と安永照雄君(大阪大学遺伝子情報実験センター教授)とで解析してみると、2つのタンパク質を作るからといって変化に対する制約が強いということはないことがわかり、オーバーラップ遺伝子も変化できることを理論的に証明できました。これが生物学での最初の論文で、1978年のネイチャーに掲載されました。私の結果は、DNAが中立的に進化することをサポートするデータだったので、この論文が出るとすぐに木村資生先生(当時:遺伝学研究所/中立説の提唱者)から電話がありました。引用文献に木村先生の中立説の論文を引いていなかったのです。物理から来たから何も知らなかったのですよね、本当に(笑)。しかし、木村先生は「中立的に解釈してくれてありがとう」と御自身の論文を送って下さいました。「本当はこれを引用するんだよ」という意味だったのでしょう。これがきっかけで、その後木村先生からご指導いただくようになりました。
その後も塩基配列の比較解析を続けました。本庶佑先生(京都大学大学院医学研究科教授)との共同研究で、マウスの免疫グロブリンγ1とγ2b遺伝子が進化の過程で相互に部分的に配列を交換しながら進化していることを示しました。これは高等動物で遺伝子変換を示した最初の例となりました。この研究で、初めて自分が分子レベルで生命現象を研究しているのだなという実感を持てましたね。

九州大学時代、北海道の学会で。左:五條堀孝(遺伝研・教授)・中央:本人・右:安永照雄(大阪大学・教授)。

最初の学生だった五條堀君と。
(左:本人 右:五條堀教授)。
中立説の検証
1980年にアメリカの遺伝学の大家クロー先生が研究室にいらっしゃいました。その時に、彼と同じ学科のスミーシスがαグロビンの遺伝子に配列がよく似ている、しかし途中に翻訳の終わりを意味する終止コドンが入っている変な遺伝子を見つけた、という話をしてくださいました。当時は遺伝子の塩基配列がわかると論文になる時代。私はその時すぐに、これは中立説の検証になると思ったので、クロー先生に「その遺伝子に機能はあるのか」とだけ質問し、「ないだろうと考えている」という返事でした。機能がないとしたら、中立説では進化速度が最大になるはずです。一方、自然選択の立場で考えると、機能がないので選択にかからないわけですから、進化速度は0になるはずです。すなわち、どちらの説を採るかで正反対の結果が期待されることになります。これは面白いことに気がついたと思い、すぐに計算して、1週間で私と安永君の連名で論文を仕上げました。結果は進化速度が最大になり、中立説をサポートする強力なデータとなりました。

折りに触れ頂いた木村先生からの便りとサインとコメントが入った中立説の本。
ホモロジーサーチ法の開発
1980年に入ると、さまざまな遺伝子の配列がどんどん分かってきました。ある時、異なるリン酸化酵素遺伝子の配列比較から、リン酸化という共通の機能を持つ遺伝子の間では、配列が良く似ているという報告が出ました。私はこれを逆側から受けとめました。つまり配列がよく似ているなら、それらの機能がよく似ているのではないか思ったのです。良く似ている配列を検索することで機能を推測できるのではかということです。このアイデアで、当時私たちが自前で持っていた3000ほどの遺伝子配列を含むデータベースを使ってホモロジーサーチのためのコンピュータシステムを作りました。
この方法を使って、1983年に当時大学院生だった藤博幸君(京都大学化学研究所バイオインフォマティックスセンター教授)、林田秀宜君(奈良県立医科大学助教授)と私はRNAを鋳型としてDNAを合成する逆転写酵素の配列を肝炎ウイルスとモザイクウイルスに発見しました。こうして機能が分かっていない塩基配列と類似の配列を持つ遺伝子を探し出すホモロジーサーチの方法が確立できたのです。実は、ラッセル・ドリトル博士が全く独立に同じ方法に辿り着いていました。しかも彼らの方が早かったのです。この後は、がん遺伝子の検索などの依頼が次々とあって、この方法で仕事をしていたのですが、86年頃、材料を変えているだけの銅鉄主義に飽きてきて、新しい問題を思いついたこともあって、ホモロジーサーチから離れようと考えるようになりました。どんどんデータは出て、よい雑誌には出たんですけどね。
オス支配進化説の提唱
ズッカーカンドルとポーリングは、分子時計(進化の過程で塩基の置換が時間に比例して起こること)を発見しましたが、私はずっとこのことを考えていました。分子時計の発見は後の分子進化学の発展に大きな影響を及ぼしました。分子系統学と中立説の誕生に影響を与えたのです。この二つは分子進化学の2つの大きな研究の流れとなりました。ところで分子時計はとても不思議な性質で、塩基置換という生物的な現象が物理的な時間(普通の時間)に比例して起こり、世代時間という生物的な時間には比例しないのです。今では多くの研究者は分子時計の一般性を信じていません。分子時計を理解しようとしているうちに、進化に寄与する突然変異の由来を考え始めていました。そして、DNAの複製エラーがその主要因であろうという結論に達しました。
そうだとすると、DNAの複製エラーは細胞の分裂の際に生じるはずですから、進化に寄与する突然変異率は生殖細胞の分裂数に比例するはずです。ところで、定量的な差は分かりませんが、卵と精子では分裂回数がずいぶん違っていて、精子の方が卵よりずっと多いと思われます。そうすると、オスの方がメスよりDNAの複製の回数、従ってDNAの複製エラーが圧倒的に多いことになります。こうして、DNAの複製エラーが進化に寄与する突然変異の主要因と仮定すると、オスの方がより多く突然変異を生成しているのではないかということに気がつきました。中立説では進化速度は突然変異率に比例しますから、オスが進化を決めるという仮説が成立します。銅鉄主義もありましたが、この問題がやりたくてホモロジーサーチが疎遠になったというのが本当のところでしょう。ずっと後でクロー先生が教えてくださいましたが、突然変異がオスに支配されているという考えは、ハーディー・ワインベルグの法則で有名なワインベルグが1910年代に既に気がついていたようです。また、1930年代には集団遺伝学のホールデンも気が付いていたらしい。
幸運にも私は大事なことに気がつきました。すなわち、精子と卵の分裂数に差があると(精子の分裂数/卵の分裂数=αとする、普通α>1)、常染色体と性染色体X、および Yの間で突然変異率が違ってくることを簡単な数学で示し、オス支配進化説(Male-driven Evolution Theory)が出来上がりました。これによってこの問題を分子生物学に乗せることに成功したわけです。例えば、ある常染色体がメスを経由するかオスを経由するかは1/2の確率ですが、Y染色体は常にオスを経由します。精子の分裂数が卵に比べて圧倒的に大きければ、オスを経由したときに突然変異が生じることになるので、この条件ではY染色体は常染色体に比べ、2倍の突然変異率を持つことになります。突然変異率はαで表され、哺乳類のようにオスがヘテロな性染色体を持つXYオス/XXメス系では、突然変異率はY染色体>常染色体>X染色体の大きさの順になります。
面白いことに、トリでは哺乳類とは逆にメスがヘテロに性染色体を持ちます。このことから鳥類では突然変異率が哺乳類の場合と逆転して、X染色体>常染色体>Y染色体の大きさの順になります。このことはこの理論の正否を検証する上で重要な性質です。
私と林田君、それに隈君(京都大学化学研究所バイオインフォマティックスセンター助教授)は常染色体、X染色体及びY染色体にある遺伝子について、ヒトとマウスで比べ、アミノ酸を変えないDNAの変化(同義置換)の速度を計算しました。すると、理論の予想通りY染色体が速く、Xが遅くなって、オス支配進化説を哺乳類で検証できました。ただ鳥類の解析はこの時点ではできませんでした。この理論と哺乳類の解析結果をコールドスプリングハーバー・シンポジウムで話しました。講演が終わるとワトソン博士(2重らせんの発見者)が近づいてきて、とても面白かったと言ってくれました。本当に嬉しかったですね。この理論はその後、ネイチャー誌上で激しい議論が起こりましたが、私は参加しませんでした。この理論の鍵は鳥類での解析だからです。1997年になってやっとスウェーデンのグループによって鳥類の解析が発表され、われわれの理論が立証されました。ある時、福岡で木村先生とタクシーに乗ったときでしたが、先生は、こんな簡単な数学で面白い理論ができるなんて驚きですね。数学が簡単だからいいですね、と言ってくださったときは、正直、子供のように嬉しかったですね。
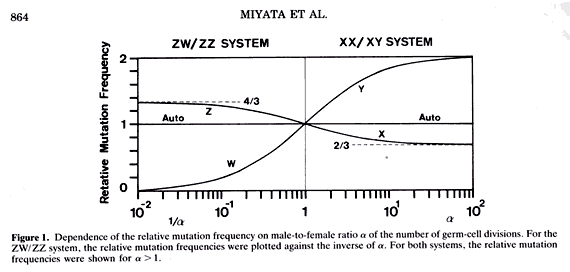
議論を巻き起こした「オス支配進化説」1987年:精子と卵の分裂数に差があるために、染色体間で突然変異率が違ってくる。哺乳類のようにオスがヘテロな性染色体を持つ場合(メス:XXオス:XY)、突然変異率はY染色体>常染色体>X染色体の大きさの順に、トリでは逆にメスがヘテロな性染色体を持つため(メス:ZWオス:ZZ)、変異率が哺乳類の場合と逆転することを示した。1997年にスウェーデンのグループによって鳥類の解析が発表され、理論が立証された。
生物最古の進化
1977年、分子進化学の一方の大きな分野となっている分子系統学(分子で生物の系統を再現する研究)で分類学上大きな発見がありました。ウース博士のグループはバクテリアを分子系統学的に分類したところ、異常な環境(たとえば100度以上の高温)で生息するバクテリアが一つの独立したグループになることを発見し、古細菌と名付けました。そして地球上に生存する全生物は、古細菌と真正細菌(普通のバクテリアのグループ)、それに真核生物を加えて、3つの超生物界からなると主張したのです。この主張はすぐに認められましたが、三者がどういう順番で分かれたのか、すなわち生物最古の進化をめぐって、10年以上の長きに亘って論争がありました。そのわけは、真正細菌、古細菌、真核生物の系統関係を知ろうとしたら、三者より遠いグループ(外群という)を入れて系統解析をする必要がありますが、今の場合、この三者で地球上に棲息する全生物になり、外群が存在しません。このために正確な系統樹が描けず、論争が続いていたのです。
実はうまい手がありました。遺伝子族の系統樹を研究していて偶然思いついたのですが、外群となる生物Xを探すと困難にぶつかりますが、遺伝子を探せば困難が回避できます。つまり、遺伝子重複で作られたポリペプチド伸長因子EF-TuとEF-Gのように、すべての生物が対で持っている重複遺伝子を使えばうまくいくことに気がついたのです。さっそくこのアイデアで岩部直之君(京都大学大学院理学研究科助手)が中心になって解析してみると、なんと異常環境に棲息している古細菌が真核生物に近縁だったのです。この研究が契機になって生物最古の進化の研究が急に進みました。少し引いた視点からみるとうまい解決法を思い付くものなのですね。
生物の多様性と遺伝子の進化
1990年に京都大学に移りました。これまでは、分子の進化機構と分子系統学をやってきましたが、そろそろ、形態レベルの進化と分子レベルの進化をどう繋ぐかという究極の問題に本格的に取り組もうと思いました。ただこのままでは実際に研究を始めることができません。こういう大きな問題に取り組むには問いの立て方が重要です。問題をうまく解けるようにほぐす必要があるのです。考えあぐねた末に、カンブリア爆発(およそ6億年前、カンブリア紀と先カンブリア時代の境で起きた多細胞動物の爆発的多様化)が起きた当時、遺伝子も連動して爆発的に多様化したか?という問題に焼き直しました。これは生物の多様性と遺伝子の多様性の関連に関する問題の一つで、これならどうやら解けそうです。
具体的には、細胞間情報伝達や形態形成に関与する多細胞動物特有の遺伝子を考えます。こういう遺伝子は進化の過程で遺伝子重複を繰り返して、たくさんのメンバーからなる遺伝子族を形成しているのが普通です。こうした遺伝子族の幾つかを分子系統学的に解析して、遺伝子重複が何時起きて新しい遺伝子が作られたのかを調べるわけです。遺伝子重複が起きた時期を知るために、系統樹の中にたくさんの動物種を含める必要があります。マウスやショウジョウバエといった実験動物についてはたくさんの遺伝子の塩基配列が決定されていますが、カイメンのような分子生物学ではめったに使われない生物の遺伝子がむしろ必要なのです。こうした生物の遺伝子の配列データはほとんどありませんので自分で用意しなければなりません。そこでついに自分で遺伝子の配列データを決めることにしたのです。研究室全員が素人だったのですが工夫しながら何とかデータが出るところまでこぎ着けました。研究室の全員で手分けしていろいろな遺伝子族の実験と解析をしました。
結果は予想していたものとはずいぶん違うものでした。遺伝子は確かに爆発的に増えていましたが、その時期は予想とは違って、およそ9億年前のカイメンとそれ以外の動物が分かれた以前に集中的に起きていました。もう一つは、脊椎動物の系統で、無顎類と有顎類が分かれたおよそ5億年前のあたりで遺伝子多様化が起きています。しかし問題のカンブリア爆発が起きた6億年前では遺伝子重複はほとんど起きていなかったのです。この遺伝子多様化パターンが意味する最も重要な点は、遺伝子の爆発的多様化はカンブリア爆発の直接の引き金ではなかったということです。カンブリア爆発と遺伝子爆発の間には時間的ずれがあるのです。このことは、カンブリア爆発の分子機構を考える上で、新しい遺伝子を作るというハードの視点ではなく、すでにある遺伝子をいかに利用してカンブリア爆発を達成したかというソフトの視点が大事なのだということを意味します。多様化のソフトモデルと呼んでいます。
カイメンとその他の動物が分岐した以前に多細胞特有の遺伝子が爆発的に増えたわけですが、それは多細胞動物に最も近いといわれている単細胞原生生物の立襟鞭毛虫から動物が分かれた後のことなのだろうか。この疑問に答えるため、菅裕君(バーゼル大学ポスドク)が中心になって立襟鞭毛虫の解析を行いました。この生物を研究している人が日本にいないため、材料の入手や、それを増やすことなど、かなり苦労しました。やっとクローニングまでこぎ着けて解析してみると、なんとまた予想が外れました。多細胞動物に特有の遺伝子が立襟鞭毛虫と多細胞動物が分かれた辺りで一斉に作られたという結果でした。多細胞でもないのに、立襟鞭毛虫には神経や細胞接着に関係する遺伝子など、まるで多細胞かと間違えるほど、多細胞特有の遺伝子が見つかってきました。多細胞生物には多細胞独特の遺伝子があって、そうした遺伝子が多細胞らしさを形作っていると信じてきたのですが、その考えはどうやら捨てなければならないようです。多細胞動物の起源を理解するために、立襟鞭毛虫のゲノム計画は面白いと思いますね。今私の研究室の岩部直之博士が計画中です。
多様化のソフトモデルを検証するため、大阪大学微生物研究所の岡田雅人教授の研究室では立襟鞭毛虫やカイメンの遺伝子をマウスの培養細胞に導入する実験をしています。それから擬態はソフトモデルそのものではないかと考えています。昔から擬態にはとても興味があって、調べてみたいと思っています。私は今年定年ですけど、なんとか研究に関わっていきたいと思っています。擬態の分子生物学が展開できるといいなと夢見ています。

木村資生賞受賞の際に。

5章の写真のメンバーの30年後!?
左:五條堀教授・中央:本人・右:安永教授。

銀閣寺の前で、遊びに来たSvante Paabo博士と。
御乱心!?
一応、京大での勤めを終えることになり、やり残したことはたくさんありますし、これからも関われるものがあれば関わっていきたいと思っています。自分ではたいしたことはできなかったなと思いますが、そういうことはあきらめます。これは私の考えですが、私がどくことで、新しい自由な発想を持った若い人に研究の場が提供できるでしょう。私は理論だから、時間を贅沢に使いたいと思ってきましたが、教授職はそういうわけにもいかなくてね。これからは、贅沢に時間を使って、ヒトがもっている普遍的な性質を進化との関連で考えていきたいと思っています。信仰を持つということも進化の過程で作られているかもしれませんからね。いやぁ、こんなことを突然言うと、御乱心と言われちゃいそうですが、そうではありませんよ。真面目です(笑)。人間が持つさまざまな特徴がどれだけ進化の過程で獲得されたのか。今の私にはとても興味がある問題です。分子でどこまで迫れるか。それが私の夢のまた夢です。
(文責:工藤光子)



