

植物少年
幼いころ体が弱く病気がちだった私は、命というものに神秘的な関心を抱いていた。一生、命を扱うような仕事ができたらいい、子供心にそう思っていた。もちろん今から思えば、漠然としたもので、それが宗教哲学になってもおかしくはなかったろう。生物学に進んだのは、ただちょっと数学ができたからにすぎない。
国民学校を卒業したのがちょうど終戦の翌年、新制になったばかりの中学に入学して、私は理科クラブに所属した。丹波の山奥のことである。先生も理科が専門というわけではなく、調べるにしても古い牧野図鑑くらいしかないのだが、それでも野山を歩きながら名前をつけることから始めた。その当時から昆虫少年ではなく、なぜか植物だった。
そんなことをやっていたものだから、高校へ入学すると、生物班に勧誘された。私としては、博物的なものに惹かれたのではなく、命とは何かということを調べたかったから、顕微鏡で微生物をのぞけるという誘い文句に惹かれて入部した。入ってみるとガの幼虫図鑑を出した先生や、地方の植物誌を精力的に調べている先生などがおられた。後に探検家になった先輩もいて、虫のこと、クモのこと、動物のことなど熱心にやっていた。田舎の高等学校では、男子が動物で女子が植物というのが慣例のようだったが、私はここでも自然に植物を担当することになった。そのころ、ホリデー・サイエンティストでナチュラリストの稲田叉男さんというシダに詳しい人がいて、山を一緒に歩いたり資料を送ってくださったり、手弁当でいろいろ教えてくださった。
入学した京都大学理学部には、シダの分類が専門の田川基二先生がいらっしゃった。田川先生のところには、学外からもナチュラリストがたくさん集まってきていた。その一つが「シダ類談話会」で、稲田さんもそのメンバーだった。私も誘われて参加し、大学とはまた違う田川先生の講義を聴き、終わった後のビールを楽しむようになった。まだ専攻は決まっていなかったが、京大の学生でこんなところに来るくらいだから、きっと自分のところに進学するだろうと期待しておられたようだった。なにしろ分類を専攻する学生は、そうそうはいない。ある日、「将来何をやるのだ?」と聞かれ、「命を解明するためには物質レベルで詰めていかなければならない。生物化学をやろうと思っている。」と、今から思えばずいぶん不遜な答え方をした。それでも期待に応えられないのをカバーするつもりで、「分類学は面白くなってきたので、一生趣味ではやっていこうと思う」などと、ずいぶん失礼なことを言ったものだった。
田川先生はあまりものを書いたりせず、教授になったのも定年前の1年半だけで、学界に対して影響力の強い方ではなかった。しかし、分類学を科学にするにはどうしたらいいかということを非常に真剣に考えている方だった。私は先生の問いにあんな答え方をしておきながら、山を一緒に歩いたり毎月の談話会に参加して話を聞いたりしたが、それは私のその後に大きな影響を与えたと思う。
私が入学した1953年は、ワトソンとクリックがDNAモデルを発表した年である。時代は分子生物学に向かって流れていて、分類学はすでに過去のものだという印象が強く、私も趣味でやるものだとしか思っていなかった。しかし田川先生の話を聞いているうちに、生きているとはどういうことかを明らかにするうえで、さまざまな生き物の生きざまを知ることが、どうしても必要だと思うようになった。しかも、それはほとんどまだわかっていないのだ…。そろそろ1年が過ぎようとするころ、「生物学に対する自分の考え方が間違っていました」と、改めて先生にお願いにいった。本当に植物分類学をやることに決めたのは、そのときである。

高校時代。生物班の海浜実習で天の橋立を訪れた。

大型シダ・リュウビンタイの調査のため紀州に行った、大学4年のときのスナップ。

博士課程4年、研究室で。このころ、こっそり童話を書いていた。

コケシノブ

ヒメシダ

ホウビシダ
シダとの馴れ初め
分類学のなかでもとくにシダを扱う気になったのは、エドウィン・コープランド(フィリピン大学農学部の創設者)の影響だったかもしれない。彼は、カリフォルニアの乾燥地帯からミンダナオ島の湿潤地にやってきて植物の多様性に驚き、シダ植物を材料にして、生態的多様度の研究をした人である。コープランドが一貫して言っていることは、生物の体系を作るときに、しばしばわかりやすさや便利さによって類型化することが優先され、せっかくさまざまな生物を扱いながら、生物相互の間にあるナチュラルな系統関係を純粋に反映する体系になっていないということだった。種子植物の場合、花が非常に有効な指標になるので、類型的にまとまりやすい。それに対してシダ植物は生殖器官の形質が限られているため、どうしても栄養器官でシステムを論じることになる。類型化はしにくい反面、逆に、本当に自然な系統関係を念頭において追及するのが日常的になる、と論じた。
コープランドをもっとよく知りたいと思った私は、学部生のころから、コープランドの扱っていたコケシノブを勉強し始め、マスター論文にまとめた。植物はふつう、基本組織系と維管束系と表皮組織系の3つの組織系からなり、単純化が進んで2層の組織を作ることはあるが、コケシノブの葉のように細胞層1層までいくのは非常に特殊な飛躍があったと考えられた。しかもこの1層は、基本組織の性質も表皮組織の性質も合わせもっている。この謎を追いかけているうちに、シダとのつき合いがますます深まることになった。
もう一つのきっかけは、陸上植物がどういうふうに陸に上がってきたかということについての関心である。現生生物の多様性は30数億年の生命の歴史の反映であるから、現生のものとの対比から、何がどこまでいえるかを検証してみたいと思ったのである。陸上植物の起源の問題を考えるうえで、シダは絶好の材料である。


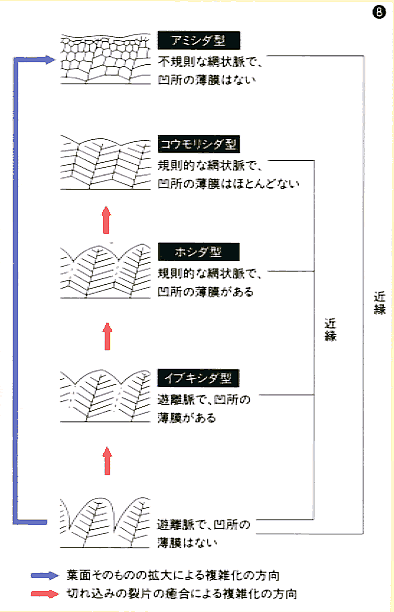
脈理の型から推定されるヒメシダ科植物の類縁関係
シダも高等植物も葉の模様(脈理)は、葉面積を拡大する方向に進化している。従来、ヒメシダ科植物は、ヒメシダ型遊離脈からホシダ型網状脈を経て、アミシダ型網状脈に進化すると考えられていた。岩槻博士は若いころ、凹所の薄膜が葉縁のニカワ質と同じ細胞構造をもつことに着目し、この薄膜をもつものは、切れ込みの裂片の癒合によって複雑化したのだろうと考えた。その結果、アミシダ型はホシダ型とは別の道すじを通って進化したと推定した(図は1962年の論文より改変)。
無融合生殖をするシダ
コケシノブの組織を追ううちに、ホウビシダが研究対象になってきた。ホウビシダには、有性生殖種と無性生殖種があることもわかってきた。ちょうどそのころ私は京大から東大に移ったばかりで、新しく担当した学生とホウビシダの生殖をテーマに調べることにした。
高等学校の教科書には、シダ植物には2倍体の無性世代と半数体の有性世代があって、規則正しく世代交代するのだと書いてある。ところが、無融合生殖種では、胞子を形成するときに減数分裂をせず、造卵器の中で卵子を、造精器の中で精子を作って受精するはずの有性生殖が完全に省略されてしまって、単為的に胞子体が形成されるのである。
高等生物は30数億年かけて、有性生殖といううまいやり方で変異をその集団内に拡散しながら進化してきた。有性生殖はものすごいエネルギーを必要とし、いうなれば相当の無駄をやることになるのだが、それにもかかわらず、生物が多様性を維持し発展させるためにはそれが一番有効な手段である、という進化の仕方をしてきた。
有性生殖をやめるのは、いったい得なのか損なのか。
無融合生殖種は、突然変異で生じたものを集団の中に拡散することができないから、種内変異が乏しくなるはずである。ところが実際には、無融合生殖をするシダの多くのものには、かなりの変異が見られる。偶然、無性生殖型のホウビシダの配偶体に造精器をもっているものが見つかった。この精子が有性生殖種と交雑して、有性生殖種の変異を集団の中に取り込んでいるのではないか…。実際にそうしていることが、アイソザイム多型を指標にして変異の起源をたどる遺伝解析によって証明され、話題を提供することになった。このことは一足先にアメリカでも、ペラエアという種について報告されていた。
ヤブソテツやベニシダ、イタチシダも無融合生殖種が多く出るグループなのだが、いずれの場合もその元になるはずの有性生殖型は、人里離れた自然環境に局限されている。そう思って探したところ、ホウビシダの有性生殖型も中国の峨眉(がび)山の山中で見つかった。また、オオヒメワラビ、オニヤブソテツなどの無融合生殖型だけが知られていた種にも、四川や雲南では有性生殖型のあることがわかってきた。
原始自然の状態では、多少エネルギーの負担が大きくても有性生殖を行っていた種が、環境が急激に変化したときに、その新しい場所に飛び込むためには有性生殖をとおしての進化では間に合わない。しかし、単為生殖をしていたのでは種として進化する力はない。何か簡単でいい方法はないかと工夫したのが、造精器をもつタイプの無融合生殖だったのではないだろうか。
日本は外国に比べて、無融合生殖種の率が非常に高い。一見すると、生物の多様化が大きく進んでいるようにみえるが、進化の流れのなかでみると、やはり特殊である。弥生時代以降、生活域が人為的に大きく撹乱されたために、植物もこれに緊急対応して生き延びてきたということを示しているのだと推測される。

インドネシアの植物園会議にて。左は英国王立キュー植物園長のプランス卿(1992年)。

国際植物園連合総会で会長に選出され、交換文書にサインしているところ(1993年)。

ライデン大学の伝統ある学位審査会に、審問官の一人として招かれた(1977年)。
パンドラの箱
私は、科学的好奇心に忠実に、今何が一番おもしろいテーマで、生物学の世界で何が解明される必要があるかを考えて研究してきた。しかし、その過程で知りえたことから、地球上で危険なことが起こりそうだというサインを読み取ったら、それを世の中に訴えていくのが、パンドラの箱を開けた科学者の義務だと思っている。
東大付属小石川植物園の園長に就任した私の仕事の一つは、絶滅の危機にある種を知り、それを象徴にして自然界に生物多様性の保全の方策を立てることだった。赴任して間もなく、小笠原諸島の父島にたった1株だけ生育している、まさに絶滅寸前のムニンノボタンを救う試みが、下園文雄さん(東大植物園技官)の熱意を中心になされた。植物園で株を増殖させ、さらに小笠原の自生地に植え戻すことに成功したのである。しかし、これらは同じ株から増殖したもので、すべて同じ遺伝子をもっており、種としての遺伝的多様性が生じるまでに気の遠くなる間、人為的な管理を必要とするだろう。本来の野生種とは、もはやいえまい。
「生きている」ということは、個体が生きているメカニズムを発揮するだけでなく、他とのかかわりのなかにあるということでもある。生物多様性を守る意味もそこにある。どんな種でも、ひとたび失われたならば、人間を含めた他のすべての生き物の生きている状況を変えてしまうのである。


分類学の夢
振り返ってみると、分類学を専門とするようになったのは、決定的なきっかけやドラマティックなできごとがあったというわけではない。さまざまな人との出会いや周りの状況があって、自然にそうなってきた。
そうした人との出会いのなかで、これからの生物学や分類学のことなどを真剣に議論したものだが、あるとき、私は二つの夢について語ったことがあった。
一つは、遺伝子を使って論じられるような分類学をやりたいということだった。形質をみる場合でも、その器官が形成される過程を追っていくことで形質の本来の姿に迫っていく。最終的には、遺伝的な情報がいかに変換されて、その変換されたものが生物の形質にどう反映され、さらに、その形質が自然の環境とどうマッチして進化してきたか。生物のもっている38億年の歴史が多様性のうえにどう反映されているか。それを見ていくのが、生物学的に正しい系統分類体系なのだと思う。今では、DNAを指標にして系統を追跡することも可能になってきた。生物多様性を普遍的な生物現象として詰めることができるようになったのである。
もう一つの夢は、ボルネオやスマトラなどの非常に多様度の高いところで思いきりフィールドワークをやりたいということだった。当時は、国内旅行もままならず、米と標本用の古新聞をもっていかなければならなかった時代である。外国へ調査に行くなど夢のまた夢だった。今でも、何千万種といわれる地球上の生物は、まだ150万種しか調べられていない。汗を流し泥だらけになって、山へ行って未知のものをとってくるということから始めなければならない。そういうレベルの仕事があって、生物の進化の全体像が詰められる。生きているとはどういうことかが本当にわかるのは、そのような調査研究の積み上げがあってのことなのだと思う。
自分のやっていることは、富士山のてっぺんから見たら1合目くらいのことかもしれないが、つねに頂上を目指しながら歩くのだと話したのは、大学院に入ったころだったろうか。この二つの大きな夢は――部分的にはかなえられているものの――今でも私は指向性としては貫いている。
