年度別活動報告
年度別活動報告書:2003年度
脊椎動物の頭部神経系はどのように部域化されるのか
橋本主税(主任研究員)
辻咲織(大学院生)
山口真未(大学院生)
重永幸(大学院生)
緒方友香(大学院生)
外谷雅範(大学院生)
皐裕美(研究補助員)
はじめに
脊椎動物の頭部神経組織は元来皮膚となりうる組織である。この組織がオーガナイザーと呼ばれる機能領域の働きにより、大まかに部域化された神経組織へと誘導され1)、それが発生過程を追うに連れて厳密に領域化が進み、最終的には隣り合う個々の細胞がそれぞれに特有の働きを担うまでに特殊化(領域化)される。このように元々均一な組織に厳密な領域情報を与える仕組みは、いわば緩やかな位置情報からシャープな位置情報への転換であり、「大きな目盛りの物差し」によって「細かな構造」を正確に形成する仕組みである。これは脳の形成に限らず生きものの形づくりの過程で頻繁に利用されている重要な方法論であるが、その詳細はまったく分かっていないため、生きもの一般に存在する秩序の形成を考える上で非常に興味深い2)。また、脳の高次構造自体が脳の高次機能を直接的に担うことが知られていることから、脳が形成される仕組みの解明が脳機能の解明へのひとつの方法論となりうる。
脊椎動物の頭部神経系の構造は種を越えて高度に保存されており、またその構造を作り出す仕組みも共通の分子機構によって担われているが3)、実際の頭部神経系を見ると、その大きさや形などが大きく異なった結果、異なる種として存在するわけで、このような種間における類似性と相違性を生み出す仕組みを知ることも、構造と機能の進化を知るアプローチとなりうる。
我々は、アフリカツメガエル(以下ツメガエル)を実験材料として形態学と分子生物学的手法の両面から研究を遂行している。ツメガエルの頭部神経形成過程では、部域化されているオーガナイザー組織が予定頭部神経組織に接し、自己の持つ位置情報を与えることで領域化された神経を誘導する4)。この領域化の過程においてもっとも興味深い点は、オーガナイザーがいかにして厳密に領域化されるのか?オーガナイザーから出る領域特異的シグナルの正体は?また、そのシグナルを受けた頭部神経はどのように振る舞うのか?である。これらの全体像を明らかにすることで、脳の形と働きの解明は言うに及ばず、生きものの高次構造形成に潜在する普遍的な真理に到達できるのではないかと期待できる。
結果と考察
1)オーガナイザーの部域化
オーガナイザーは、最終的に咽頭内胚葉・内中胚葉・脊索前板・脊索などに分かれる。(図1)このオーガナイザーの領域が神経系の領域形成に決定的に重要であることが知られているが、この様な詳細な領域化が初期のオーガナイザーにおいて成立しているかについては現時点では明らかではない。また、オーガナイザー由来の各領域の神経組織の領域化に対する働きに関しても諸説あり、どのオーガナイザー領域がどの神経領域の形成に機能しているのか定かではない。そこで我々は、予定脊索前板に発現する遺伝子Xhairy2b5)に着目して、脊索前板がオーガナイザー内部に確立する時期を探り、さらに脊索前板が頭部神経のどの領域の形成に関与するのかについて解析を行なった。(図2)
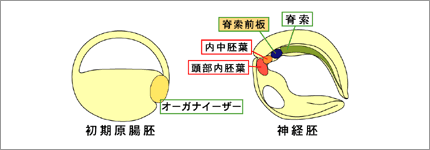
図1:オーガナイザーは発生が進むにつれ複数の領域に分かれる
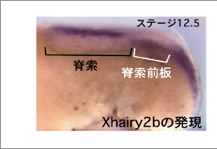
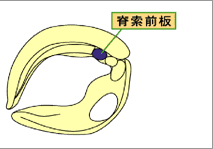
図2:Xhairy2bは脊索前板で発現する
まず、Xhairy2b遺伝子はそれ自体が体幹部を誘導できることが明らかとなった。この事実は、現在までに提示されている「脊索前板は頭部誘導能を持つ」という説とは異なり6)、「脊索前板は体幹部の誘導に働く」ことを示す。また、Xhairy2b遺伝子は脊索前板特異的な遺伝子群の発現を正のフィードバック制御によって互いに誘導できることを見いだした。この事実は、初期オーガナイザー領域内部でXhairy2b遺伝子の発現が起こる場所では必然的に脊索前板特異的遺伝子の発現が継続的に維持されることを示す。さらに、脊索前板に隣接する他のオーガナイザー領域である内胚葉や脊索に特異的な遺伝子の発現がXhairy2bによって抑制されることが明らかとなった。これらの事実を統合すると、オーガナイザー内部の特定の細胞に発現したXhairy2bは、その細胞では脊索前板特異的遺伝子の発現が保証するとともに、隣接する組織への分化転換を積極的に抑制することにより、予定脊索前板のアイデンティティーの確立に重要な働きをしていると考えられる。(図3)
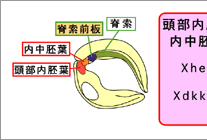
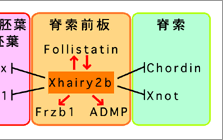
図3:Xhairy2bによる遺伝子発表制御
さらに予備実験として、オーガナイザーにおけるXhairy2b遺伝子の機能を人為的に阻害したところ、脊索前板の形成が抑制され、また頭部神経系の特定領域の形成に影響が見られた。これらの結果から、まだ形態学的に各領域が確立していない初期オーガナイザーにおいて、少なくとも予定脊索前板領域は潜在的に成立しており、それが頭部神経系の特定領域の形成に重要な役割を担う可能性が示された。これは、脊索前板が体幹部誘導能を持つと同時に脳の特定領域の形成にも関与することを示す。頭部誘導能を有するが体幹部を誘導できない内胚葉領域と、体幹部誘導能を有するが頭部を誘導できない脊索に挟まれて存在する「脊索前板」の特異な機能のひとつをかいま見ているのかも知れない。
2)オーガナイザーから頭部神経組織へのシグナル
オーガナイザーはそれ自身が領域化され、各オーガナイザー領域からの位置情報が頭部神経組織の位置情報となる。現在までに我々は、転写補助因子Groucho依存的に予定間脳領域の背腹が決定されること、そしてその領域化を制御するシグナルが初期オーガナイザーから生じることを見いだしている。Grouchoは頭部神経組織の細胞内で機能するが、オーガナイザーから頭部神経へのシグナルは細胞間の情報であることから、オーガナイザーの特定領域から出された細胞外分子が予定間脳領域に働きかけ、その結果としてGrouchoを介した細胞の運命決定が行なわれると想定し、頭部神経細胞内でGrouchoと相互作用する転写因子の特定と、オーガナイザーから出される細胞外分子の特定の両方向からアプローチをかけた。第一に、初期オーガナイザーが形成される時期に神経細胞特異的に発現している転写因子はほとんど知られていないため、Xotx2・Sox2など既知の分子に狙いを定めて、その転写促進型変異体および転写抑制型変異体を作成して、間脳の背腹軸に及ぼす効果を調べた。結果として、解析した全ての転写因子に関して間脳背腹軸形成を制御しうるものは確認できなかった。次に、標的細胞内でGrouchoを介して転写制御を行なうことが知られている成長因子Wntに着目して解析を行なった。Wntシグナル経路では、最終的な転写因子であるLEF/Tcfに普段はGrouchoが結合して転写を積極的に抑制しているが、一度Wntシグナルが活性化されるとβカテニンがGrouchoに取って代わってLEF/Tcfと結合し、その標的遺伝子の転写を促進すると考えられている。Wntシグナル経路を活性化する分子としてβカテニンの他にはdishevelledが、逆に抑制する分子としてはGSK3やAxin/Axilが知られている。我々はAxin/Axilによるシグナル経路の不活性化や、βカテニンによる活性化を行ない、間脳の背腹軸決定に影響があるか解析したところ、期待通りの結果を得ることができた。次に、βカテニンあるいはGrouchoとの結合能をそれぞれ欠く変異LEF/Tcfを作成し、その効果を解析した。我々の仮説によると間脳背側領域でWntシグナルが活性化されており、間脳腹側領域ではWntシグナルは抑制されているはずであるが、βカテニンとの結合能を欠くLEF/Tcfを強制的に発現させると背側領域での活性が予想されるWntシグナルに応答できないため、結果として間脳全体の腹側化が起こり、逆にGrouchoとの結合能を欠くLEF/Tcfの存在によって間脳は全体として背側化すると期待されるのである。(図4)まだ予備実験段階ではあるが結果は期待通りであり、間脳の背腹軸決定にWntシグナルが何らかの機能を有することが示された。これは、Grouchoの機能をWRPWペプチドによって特異的に阻害したときの結果とも一致する。これから重要なことは、Wntを特異的に発現するオーガナイザー領域の同定を行ない、オーガナイザーが頭部神経の領域を決める仕組みの全体像を明確にすることである。
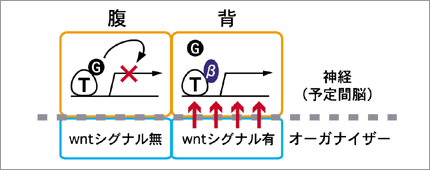
図4:予定間脳領域の背腹はWntシグナルによって決定される
(T:転写因子、G:Groucho、β:β-カテニン)
おわりに
ひとたび特定の運命に決定された細胞がその運命をどのように維持するのかという課題には、Xhairy2bの働きによってひとつのモデルを提唱できたと考えている。しかし、全く同じ環境にある隣り合う細胞の内の一方が脊索になり他方が脊索前板になるとき、それぞれの細胞運命を決める分子機構はどのようなものなのか?機械論的ではなく、確立論的にさえ見える個々の細胞の振る舞いを分子によって解き明かすことはこれからの発生学において重要な課題のひとつであり7)、本研究の最終目的のひとつにこの課題への解答があることは間違いのないことであるが、現時点ではそのヒントすら得られていない。これは「いい加減さ」や「大ざっぱさ」を許容できる「しなやかさ」を意味し、決して機械には真似のできない生きものが生きものとして存在する本質的現象である。
また、Grouchoからのアプローチにより、間脳の背腹を決めるシグナルの一端が垣間見えてきたが、あくまでも「間脳の背腹」の決定機構に過ぎない。なぜ間脳以外の領域の背腹はGrouchoによって影響を受けないのか?あるいはそもそも間脳を決める機構は何か?について解答がもたらされなければ脳の領域化を理解することはできない。まだまだスタートラインに立ったところではあるが、いままで奥の見えなかった暗闇に一筋の光が差し込んだことは事実であろう。
最後に、Grouchoの機能解析から、脳形成とは直接関わりはないが非常に重要な生命現象を現在までに見いだしている。それはGrouchoが基本転写因子と相互作用して一般的な遺伝子発現を制御している可能性を強く示唆する結果であり、今までの常識では説明のできない現象である。これをさらに解析することにより、初期発生期における遺伝子発現制御の新しい仕組みが見いだせることが期待される。
参考文献
1) Spemann, H. and Mangold, H. (1924)
Wilhelm Roux's Arch. 100: 599-638
2) Holloway, D.M., Reinitz, J., Spirov, A. and Vanario-Alonso, C.E. (2002)
Sharp borders from fuzzy gradients. Trends Genet. 18: 385-387
3) Holley, S.A. (1997)
Fish are like flies are like frogs: conservation of dorsal-ventral patterning mechanisms. BioEssays 19: 281-284
4) 4) Koide, T., Umesono, K and Hashimoto, C (2002)
When does the endomesoderm meet the anterior-most neuroectoderm during Xenopus gastrulation? Int. J. Dev. Biol. 46: 777-783
5) Tsuji, S., Cho, K.W.Y. and Hashimoto, C. (2003)
Expression pattern of a basic helix-loop-helix transcription factor Xhairy2b during Xenopus laevis development. Dev Genes Evol. 213: 407-411
6) Shibata M., Itoh M., Ohmori S.Y., Shinga J. and Taira M. (2001)
Systematic screening and expression analysis of the head organizer genes in Xenopus embryos. Dev Biol. 239:241-56.
7) Ko, M.S.H., Nakauchi, H. and Takahashi, N. (1990)
EMBO J. 9: 2835-2842